Чтобы проверить, что Wnt11 позитивные клетки субэктодермального пространства у HH22-23 эмбрионов кур предварительно отсоединились от DML и мигрировали в направлении срединной линии, сомиты трансплантировали слева направо в обратном порядке вдоль медиолатеральной оси (сомиты VIII - XII у HH15-16 эмбрионов кур) (Olivera-Martinez et al ., 2002). Др. словми, вырезали участок в 5 сомитов (Fig. 4A) на левой стороне эмбриона донора на стадии и на уровне вдоль передне-задней оси, соответствующим экспрессии Wnt-11, индуцируемой с помощью Wnt-1 из нервной трубки, но где изолированные Wnt-11 клетки ещё не присутствовали. Затем трансплантировали этот кусочек на правую сторону эмбриона хозяина той же стадии. Следовательно, медиальный сомит примыкал к промежуточной и латеральной пластинке мезодермы. Спустя 6 ч после трансплантации DML, теперь в латеральном пложении, всё ещё экспрессировала Wnt11, демонстрируя, что экспрессия Wnt11 становится независимой от Wnt1 нервной трубки. Спустя 10 ч, Wnt11 позитивные клетки мезенхимы обнаруживаются между центральным дермомиотомом и эктодермой, а спустя 24 ч они колонизируют субэктодермальное пространство от трансплантированной DML в направлении срединной линии через весь дермомиотом (Fig. 4 B,C). Это указывает на то, что экспрессия гена не м. рассматриваться как трассер клона, поэтому сейчаст ведутся экспреименты с использованием мечения медиальных частей дермомиотома с помощью RCAS-GFP маркёра..
Wnt11, однако, экспрессируется преимущественно в клетках, которые подвергаются эпителиально-мезенхимному переходу, чтобы приобрести способность к миграции. Его экспрессия поддерживается во время миграции мезодермальных клеток, покидающих первичную полоску у рыбок данио и Xenopus (Heisenberg et al ., 2000 Tada and Smith, 2000). Поэтому очень возможно, что Wnt11 позитивные клетки в субэктодермальной мезенхиме происходят из DML. У позвоночных
amniotes, мы полагаем, что Wnt-11 экспрессируется в дорсальном дермальном клоне до тех пор, пока эти клетки движутся: первые в очереди достигают субэктодермального пространства и затем верхнего плотного дермиса, который при этом реорганизуется, чтобы сформировать дермальные конденсаты и дермис между зачатками (Viallet et al ., 1998; F. Michon unpublished data). Важность Wnt семейства генов для морфогенеза кожи демонстрируется работой лаб. Elaine Fuchs (Das Gupta et al ., 2002). У эмбрионов кур Wnt семейство генов играет фундаментальную роль в организации дермы (Chang et al ., 2004), как показывают экспреименты с использованием Wnt ингибитора Dickkopf-1 (Dkk-1). Если диссоциированные дермальные клетки эмбрионов кур трансдуцируются с помощью RCAS- Dkk-1 и затем реагрегируются и покрываются сверху эпидермисом, то образование перьев ингибируется. Dkk, однако, не далает различий между разными Wnts, а специфические антогонисты для каждого из членов этого большого семейства ещё не обнаружены. Существуют очевидные противоречия между 4 разными группами в отношении наиболее медиальной части мезенхимы, которая выше нервной трубки. Проедполагается, что она не дермомиотомного, а склеротомного происхождения и д. , следовательно, давать дорсальные части позвонков (Ben-Yair et al ., 2003). Доказано, что субэктодермальная мезенхима срединной линии на ст. E4 у эмбрионров кур содержит и хрящевые, экспрессирующие Msx1, и дермальные предшественники (Monsoro-Burq et al ., 1994; 1996). У мышей тонкая медиодорсальная мезенхима, экспрессирующая Msx1 на E12, как полагают, составляет часть дермальных предшественников (Houzelstein et al ., 2000). Мы предполагаем, что происходят две разные миграции, одна из дермомиотома и др. их склеротома. Эти две миграции м. происходить в разное время. Др. возможность в том, что первичная дорсальная мезенхима м. давать две разные популяции. Фундаментально оба исследования (Olivera-Martinez et al ., 2000; 2002; Ben-Yair et al ., 2003) согласуются с тем, что медиальная часть дермомиотома генерирует большинство аксиальных предшественников дорсального дермиса. Необходимо отметить, что в торакальной области имеется не один, а lsdf инициальных рада перьев, которые разграничены semi-apterium (Dhouailly et al ., 2004). Мы полагаем, что расхождения в midline extent производных сомитов дермальных клеток, обнаруживаемы разными группами исследователей, обусловлены наблюдениями на не совсем в точности том же самом уровне и м. отражать будущий паттерн densification дермиса и м.б., таким образом, причастны к предопределению точной позиции, где дермис достаточно плотен, чтобы инициировать образование перьев.
En-1 is expressed in the central dermomyotome, then
later in the mediolateral subectodermal mesenchyme
Хотя точный механизм всё ещё неоднозначен, DML, как полагают, является источником новых клеток для роста первичного миотома и эпителия дермомиотома, за счёт образования центрального компартмента дермомиотома (Denetclaw et al ., 1997; Denetclaw and Ordahl, 2000; Denetclaw et al ., 2001; Ordahl et al ., 2001;
Venters and Ordahl, 2002). Отслеживали (Olivera-Martinez et al ., 2002) экспрессию маркёра центральной части дермомиотома En1. Наши результаты указывают на то, что вторая небольшая популяция субэктодермальных клеток, по-видимому, исходно локализуется в центральном домене дермомиотома (Fig. 5 A,C). Это наблюдение согласуется с экспериментами по DiI мечению, показавшими вклад этого центрального домена в субэктодермальную мезенхиму (Ben Yair et al .,
2003). Некоторые клетки, экспрессирующие En1, образуют слой под эктодермой и дают субэктодермальные мезенхимные клетки вследствие процесса desepithelialization, описанного (Christ and Ordahl, 1995), который м. находиться под контролем NT-3 из нервной трубки (Brill et al ., 1995). Разработан метод транскрипции En1 в присутствии хорды в качестве источника Shh, который позволяет выживать медиальным сомитным клеткам (Rong et al ., 1992; Teillet et al ., 1998, Marcelle et al ., 1999), но в отсутствие Wnt1 из дорсальной части нервной трубки (Olivera-Martinez et al ., 2001; 2002). При этом экспрессия En1 собственно инициировалась, указывая тем самым, что En1 начало не нуждается в факторе нервной трубки. Затем мы осуществляли эксцизию дорсальной эктодермы у HH12-14 эмбрионов и установили, что and экспрессия En-1 не способна активироваться на уровне эксцизии спустя 24 ч после операции (Olivera-Martinez et al ., 2002), даже учитывая, что заживление эктодермы происходит в среднем через 16 ч (наши данные и Thйvenet, 1969). Спустя 48 ч экспрессия En-1 возобновляется, но домен её экспрессии редуцирован (Olivera-Martinez et al ., 2002), при этом не выявляется аномалий в дорсальном поле перьев спустя 8 дней (Olivera-Martinez et al ., 2001). Для предупреждения контакта между эктодермой и центральной частью дермомиотома в течение длительного периода мы вставляли кусочек teflon под эктодерму на длину 3-х только что сформированных сомитов на првой стороне на ст. HH 13-14 (Fig. 5E).
Было установлено, что спустя 24 ч экспрессия En-1 отсутствует в центральной части дермомитотомов сомитов, расположенных на уровне передних конечностей, тогда как экспрессия En-1 индуцировалась на левой (контрольной) стороне, а также в последующих сомитах (Fig. 5 F,G). Хотя desepithelialisation центрального домена дермамиотома коррелирует с подавлением Wnt6 в покрывающей эктодерме (Schubert et al ., 2002; C. Marcelle, personal communication), идентичность эктодермального сигнала, который контролирует En1 всё ещё не установлена.
Какова роль En-1? Этот ген является известным репрессором транскрипции. М. предположить, что он действует как ингибитор дермальной дифференцировки, исходя из факта, что у эмбрионов кур инициации дифференцировки кожи предшествует градиент от срединной линии (где экспрессируется Wnt-11) до латерального дермиса (где экспрессируется En-1). Точнее, предермальные клетки, которые экспрессируют En-1, д.б. задержаны в своём достижении способности индуцировать перья, чему соответствует задержка у них и экспрессии Dermo-1.
Dorsal dermis differentiation in
chick and mouse embryos
Необходимо отметить, что формирование паттерна придатков кожи заметно отличается у кур и мышей. У кур первые зачатки перьев появляются в виде одиночного ряда, распроложенного по срединной линии дорсального поля перьев (т.e. в spinal pteryla), поверх нервной трубки. Затем последовательно присоединяются новые ряды в виде пар с обеих сторон от инициального ряда в виде волны дифференцировки, которая останавливается на краю дорсального поля перьев, где образуется semiglabrous
(голая) область. У мышей напротив, большая группа зачатков первичных волос появляется сочетанно сначала на боковых стронах туловища в виде будущих латеральных полей волос шерсти, а два дня спустя появляется вторая группа по срединной линии кожи (Dhouailly et al ., 2004). Различия в формировании паттерна кожных придатков коррелирует с различиями в паттерне экспрессии Dermo-1. У кур его транскрипты впервые обнаруживаются на ст. HH 24 в медиодорсальной субэктодермальной мезенхиме (Scaal et al ., 2001), где появляются и первые зачатки перьев. Напротив, у мышей Dermo-1 транскрипты впервые появляются на ст. E11 в латеральных частях туловища (Houzelstein et al ., 2000; and S. Missier, personnal comunication). На E13, экспрессия Dermo-1 выявляется во всём дорсальном верхнем дермисе (Houzelstein et al ., 2000; Li et al ., 1995), непосредсвенно перед появлением второй группы зачатков волос.
Два др. аргумента приводят к подозрению критической роли Dermo-1 в формировании дермиса. Этот ген, как полагают, (Sosic et al., 2003) д.б. назван twist-2 на базе его высокой гомологии с Drosophila twist (Wolf et al., 1991) и twist млекопитающих. Во-первых, этот ген как известно, действует как репрессор для трансактивации Myo D (Gong and Li, 2001) и на этом пути он м.б. способен стабилизировать формирование дермиса путём предупреждения развития клеток в мышечный клон. Во-вторых, twist-2 нулевые мутанты (Sosic et al., 2003) имеют тонкий рыхлый дермис на E17, а также драматическую потерю клеток за счёт обширного постнатального апоптоза, который особенно проявляется в дермисе. Этот апоптоз усиливает уже имеющийся фенотип у эмбрионов и вызывает образование редких волос.
С помощью трансплантаций мышиных сомитов хозяевам курам, было предположено, что субэктодермальная мезенхима, происходящаяя из сомитов, состоит из двух отдельных медиальной и латеральной популяций, которые экспрессируют соотв. Msx-1 и Dermo-1 (Houzelstein et al ., 2000). Те же авт. предположили, что большинство поверхностной дорсомедиальной мезенхимы подавляет Msx-1 перед переключением на Dermo-1. Кроме того, в этой работе установлено, что миграция клеток происходит из медиальной части сомита в эктодерму/дорсальную часть нервной трубки. Т.о., клетки из трансплантированного мышинй медиальной части дермомиотома способны отвечать на Wnt-1 сигнал, который исходит из нервной трубки кур и м.также управляться всё ещё неизвестным эктодермальным сигналом. Основной вопрос локализации дермальных предшественников внутри мышиного дермомиотома является , однако, всё ещё плохо документирован и ещё не изучен экспериментально. Доступны лишь немногие факты. В нашей лаб. мы установили экспрессию Wnt-11 и в медиальной части дермомиотома и затем в дорсальной мезенхиме эмбрионов мышей (see above). En-1 экспрессия была обнаружена в демомиотомк, но только в хвостовых сомитах (Davidson et al ., 1988). Sim-1 экспрессируется в латеральной части дермомиотома мышей (Ema et al ., 1996) и домен его экспрессии является большим, чем у кур. Предварительная диаграмма показывает сходство и различия у эмбрионов кур и мышей по этому вопросу (Fig. 6).
Concluding remarks
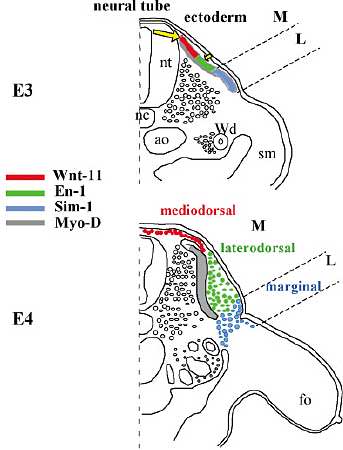
Fig. 7.
Regulation and localization of the thoracic dermal progenitors in chick embryo. At (E3) , three distinct dermomyotomal compartments express, respectively, Wnt-11 , in the most medial (activated by Wnt-1 from the dorsal neural tube (nt), En-1 in the central compartment, activated by a yet unknown signal from the ectoderm and Sim-1 in the lateral part. The Wnt-11 and En-1 expressing compartments form the medial dermomyotome. At (E4) , formation of dorsal subectodermal mesenchyme, which express Wnt-11 in the dorsomedial region, En-1 in the dorsolateral region and Sim-1 in the margin region. The latter forms the frontier with the mesenchyme originating from the somatopleural mesoderm (sm), at the intersection between the future dorsal and scapular pterylae. ao, aorta; fo, forelimb; nc, notochord; nt, neural tube; Wd, Wolffian duct. Modified from Olivera-Martinez et al. (2002). Drawing by B. Peyrusse.
У кур, как хорошо известно, имеется три домена дермомиотома, которые дают три субэктодермальных домена мезенхимы (Fig. 7). Сигнал, ведущий к экспрессии Wnt-11 в наиболее медальной части дермомиотома, четко установлен в разных лаб. как имеющий отношение к экспрессии Wnt-1 из дорсальной части нервной трубки. Скорее всего BMP4 из латеральной мезодермы ответственен за экспрессию Sim-1 в латеральной части. Функция экспрессии Wnt-11, по-видимому, связана с клеточными движениями для образования плотного дермиса, а позднее для дермальной организации и для образования дермальных конденсатов, которые ответственны за индукцию кожных придатков. Всё ещё неидентифицированные эктодермальные сигналы м. регулировать образование плотного дермиса, как это вытекает из наших исследований мутантных кур Ottawa naked (Olivera-Martinez et al ., 2004).Сегодня мы ещё не способны идентифицировать природу эктодермального сигнала, ответственного за экспрессию En-1, но диссоциация клеток, принадлежащих к центральной части дермомиотома, по-видимому, укащзывает на подавление экспрессии Wnt-6 в эктодерме (C. Marcelle, personal communication).
Что м. сказать о роли En-1 и Sim-1? Оба, как известно, действуют как репрессоры транскрипции и мы полагаем, что у эмбрионов кур En-1 м. действовать, задерживая экспрессию Wnt-11 и Dermo-1, и т. о. латеральную волну образования пллотного дермиса. Sim-1 м. выполнять и более строгую роль, т.к. densification верхней части дермиса в semi-apteria происходит с 2 дневной задержкой. Наконец, др. проблема, нуждающаяся в разрешении, это последовательность экспресисии генов в субэктодермальной мезенхиме, ведущей к образованию дорсального плотного дермиса как у кур, так и у мышей.
сеток металлического один
Сайт создан в системе
uCoz