Инструктивные (morphogen) градиенты могут формировать часть механизма, с помощью которого транскрипты онтогенетических генов, таких как Hox гены, позиционируются вдоль развивающейся оси позвоночных. Характеристика регуляторов Hox генов, которые экспрессируются в виде градиентов, может быть важной ступенью в формировании паттерна у позвоночных. Гены cdx являются вышестоящими регуляторами Hox генов (e.g., Subramanian et al., 1995; Epstein et al., 1997; Charitй et al., 1998; Isaacs et al., 1998; van den Akker et al., 2002) и cdx белки оказываются распределенными в виде posterior-to-anterior (P-A) градиентов вдоль оси гаструлирующего эмбриона (Gamer and Wright, 1993; Meyer and Gruss, 1993; Beck et al., 1995). Имеются доказательства, что за положение границ экспрессии Hox ответственны cdx белки дозово-зависимым способом и что градиенты cdx могут, следовательно, служить как instructional (morphogen) градиенты для установления паттернов экспрессии Hox (Charitй et al., 1998; Gaunt et al., 2004).
Для
cdx1, градиенты экспрессии были изучены нами ранее с использованием
cdx1/lacZ трансгенов, экспрессируемых эмбрионами трансгенных мышей (Gaunt et al., 2003; Gaunt, 2004). Мы получили доказательства, что ген
cdx1/lacZ экспрессируется главным образом в пролиферативных задних частях эмбриона и что градиенты формируются за счет распада генных продуктов в клетках нейроэктодермы и мезодермы, когда они оказываются позади регрессирующей хвостовой почки. Мы предположили, что сходный механизм может генерировать градиенты и эндогенного cdx1 белка и что формирование градиентов за счёт распада может быть важным механизмом формирования паттерна вдоль эмбриональной оси. Для проверки, приложим ли данный механизм к др. генам
caudal мы исследовали экспрессию
cdx4/lacZ и cdx2/lacZ у трансгенных эмбрионов мышей.
Discussion
Cdx/lacZ gradients formed by decay
Полученные результаты для cdx4/ и cdx2/lacZ репортерных генов подтверждают наши ранние предположения, сделанные для cdx1/lacZ относительно способа, с помощью которого образуются градиенты cdx/lacZ белков у эмбрионов мышей. Предполагалось 1) что cdx продукты образуются только или главным образом внутри задних пролиферативных зон эмбриона и 2) что клетки, покидающие эту область, подвергаются зависимому от времени распаду cdx/lacZ белка, когда они дистанцируются от регрессирующей хвостовой почки (Gaunt et al., 2003; Gaunt, 2004). Наши находки не исключают возможности, что распад cdx/lacZ мРНК также вносит хотя и значительно меньший вклад в формирование градиента. Распад мРНК обнаружен в качестве первичной причины градиента Fgf в хвостовых почках позвоночных
(Dubrulle and Pourquiй, 2004).
Используя антитела, Gamer and Wright (1993) показали, что мышиный белок cdx4 распределяется в виде P-A градиентов вдоль эмбриональной оси. Они сообщили, что природа градированного сигнала делает трудно идентифицируемыми точные передние границы, но они установили границу на 8.5 день в нейроэктодерме, которая располагалась в направлении позиции полностью закрывшейся нервной трубки (their Fig. 5D,6A) и сообщили, что cdx4 белок в мезодерме, по-видимому, распространяется, после продолжительного окрашивания (legend to their Fig. 5), в наиболее задние сомиты.
Сходным образом антитела к мышиному белку cdx2 (Beck et al., 1995) дают окрашивание, которое распространяется в направлении нервной трубки к точке полного закрытия нервной трубки. Все эти границы чётко располагаются впереди по отношению к границам мРНК cdx4 и cdx2, выявляемым в нашем исследовании гибридизации
in situ (our Fig. 5A,5N). Напротив, опубликованное полномасштабное окрашивание белка cdx4
на 8.5 день, по-видимому, сходно с нашим паттерном окрашивания cdx4/lacZ (cf. Fig
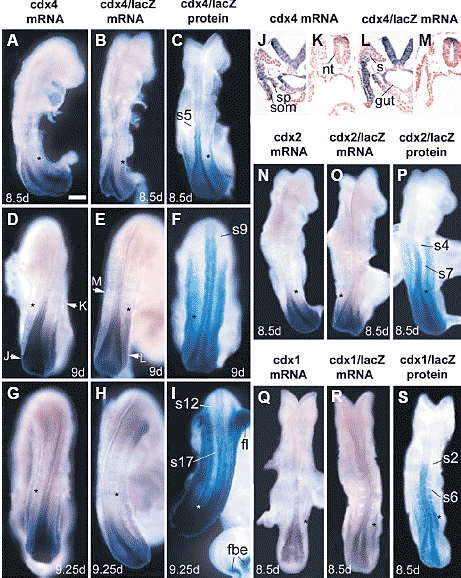 Fig. 5. Comparison of cdx mRNA, cdx/lacZ mRNA and cdx/lacZ protein distributions during gastrulation. Embryos are either wild type (A,D,G,J,K,N,Q), cdx4/lacZ transgenic (B,C,E,F,H,I,L,M), cdx2/lacZ transgenic (O,P) or cdx1/lacZ transgenic (line described in Gaunt
et al., 2003) (R,S). lacZ staining was for either 20 min (C,F) or 40 min (I,P,S). Asterisks indicate positions of the latest-formed somitic furrow in each embryo. Expression of cdx4/lacZ in forebrain and forelimbs (I; and seen, after 9 days, in all four cdx4/lacZ transgenic lines examined)
is apparently ectopic since it is not found using the cdx4 mRNA probe (G). Arrows in (D,E) indicate the positions of the transverse sections shown in (J-M). fl, forelimb bud; fbe, forebrain expression domains; other labels as for Fig. 2. Bar, 0.1 mm.
Fig. 5. Comparison of cdx mRNA, cdx/lacZ mRNA and cdx/lacZ protein distributions during gastrulation. Embryos are either wild type (A,D,G,J,K,N,Q), cdx4/lacZ transgenic (B,C,E,F,H,I,L,M), cdx2/lacZ transgenic (O,P) or cdx1/lacZ transgenic (line described in Gaunt
et al., 2003) (R,S). lacZ staining was for either 20 min (C,F) or 40 min (I,P,S). Asterisks indicate positions of the latest-formed somitic furrow in each embryo. Expression of cdx4/lacZ in forebrain and forelimbs (I; and seen, after 9 days, in all four cdx4/lacZ transgenic lines examined)
is apparently ectopic since it is not found using the cdx4 mRNA probe (G). Arrows in (D,E) indicate the positions of the transverse sections shown in (J-M). fl, forelimb bud; fbe, forebrain expression domains; other labels as for Fig. 2. Bar, 0.1 mm.
5D,E of Gamer and Wright, 1993, with our Fig 5C). Сходные заключения могут быть сделаны для эндогенного белка cdx1. Он распространяется обильно вперед, особенно заметно внутри сомитов, к позициям более передним, чем домены мРНК cdx1, выявленные в наших исследованиях, одинаково с нашим паттерном окрашивания cdx1/lacZ (cf. Fig. 4G,H of Meyer and Gruss, 1993, with our Fig 5S). Т.о., хотя cdx и cdx/lacZ белки могут быть не теми же самыми в отношении их периодов полу-жизни, но мы полагаем, что они обладают той же самой возможностью генерировать свои градированные домены экспрессии, по крайней мере частично, как результат распада белка, и lacZ окрашивание может служить чувствительным методом для выявления инициальных передних границ градиентов эндогенных cdx белков. У кур, как и у мышей, cdx мРНК, по-видимому, остаются в основном ограниченными хвостовой почкой (Morales et al., 1996; Marom et al.,
1997). У
Xenopus, однако, cdx мРНК распространяются вперед в виде градиентов вдоль нервной трубки (e.g. Pillemer et al., 1998). У этих видов, следовательно, распад мРНК может вносить больший вклад, чем у мышей или кур в формирование градиентов cdx белков.
Только механизм распада может генерировать столь гладкий градиент, если не происходит экстенсивного перемешивания клеток внутри столбов клеток, оставляющих позади себя регрессирующую хвостовую почку. Тот факт, что гладкие градиенты могут наблюдаться (e.g., Fig. 5F,P,S) доказывает, что значительного перемешивания клеток не происходит. Исследования с помощью клеточных маркеров показали прогрессивное уменьшение в A-P перемешивания клеток в развивающейся нейроэктодерме и coherent рост после 8 дня наиболее передених частей спинного мозга (Mathis and Nicholas, 2000).
Пятнистость в отношении интенсивности lacZ окрашивания, которая отмечается (e.g.,Fig. 2L) может отражать низкие уровни перемешивания клеток. Это объяснение 'salt and pepper' паттерна ранее было дано Dubrulle and
Pourquiй (2004) в отношении нерегулярностей в градиенте мРНК FGF. Это объяснение не может быть полным, однако, некоторые из наших линий и transient transgenics четко показывают значительно меньшие проявления пятнистости, чем др., как нейроэктодерме, так и мезодерме (c.f. Fig. 2E,F and 2L).
The cdx4 intron enhancer
Мы установили, что корректная экспрессия мышиного репортерного гена
cdx4/lacZ нуждается в энхансерном элементе, расположенном внутри первого интрона. Нормальная экспрессия
Xcad3, структурного гомолога
Xenopus мышиному
cdx4 (e.g. Marom et al., 1997), зависит как от вышестоящих (Reece-Hoyes et al., 2005), так и intron 1
(Haremaki et al., 2003) элементов. Критическими элементами в
Xcad3
интроне 1 являются чувствительные к Fgf элементы, которые широко распространены (Haremaki et al., 2003). Наши находки, следовательно, могут указывать на то, что наиболее важные из этих элементов для задней экспрессии мышиного гена
cdx4 расположены внутри области в 3.3 интрона, которая локализуется по направлению к его 3' концу. Действительно ли
Xcad3 является настоящим функциональным гомологом мышиного
cdx4, т.к его паттерн экспрессии, как наиболее переднего из
Xcad генов (e.g.
Pillemer et al., 1997) делает его более похожим на
cdx1.
cdx gradients and Hox expression boundaries in the developing embryo
Границы экспрессии Hox expression отвечают на cdx белки дозово-зависимым способом и градиенты cdx могут служить инструктивными
(morphogen) градиентами для закладки паттернов экспрессии Hox (Charitй et al., 1998; Gaunt et al., 2004). Для cdx1 и cdx2, кроме того, имеются доказательства из исследовании по нокауту, что оба гена вносят вклад в формирование паттерна всего вдоль позвоночного столба (van den Akker et al., 2002). Fig. 6 показывает один из способов, с помощью которого это может происходить, с помощью механизма, который комбинирует оба cdx инструктивных градиента, движущихся прогрессивно каудально по мере развития оси, и также 'temporal colinearity' в предопределении времени инициальной экспрессии Hox генов (Izpisъa-Belmonte et al., 1991). Схема может объяснить два наиболее ранних наблюдения по инициальной экспрессии Hox генов. Т.о., экспрессия Hox генов может быть градирована первой (Gaunt,
2001) , т.к. ген отвечает на градиент cdx активатора; и экспрессия Hox может существенно регрессировать кзади (Bel-Vialar et al., 2002; Gaunt et al., 2004) из-за регрессии в каудальном направлении градиента белка cdx.
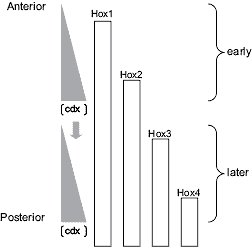 Fig. 6. Possible scheme for the establishment of Hox expression
boundaries along instructional cdx gradients. During gastrulation, the
cdx2 and cdx4 gradients regress posteriorly along the developing axis
(indicated here by a vertical arrow). At early times only 3'-located Hox genes are expressed and their boundaries may become differentially sorted by the cdx protein gradients as they then exist over prospective anterior parts of the embryo. This may occur, for example, because each Hox gene is
activated by the products of more 3' Hox genes (Hooiveld et al., 1999;
McNulty et al., 2005), but only above a threshold cdx protein concentration.At later times, more 5'-located Hox genes become expressed and their boundaries become sorted over the more posterior structures which are then displaying the cdx gradients. The protein gradients from all three cdx genes likely overlap (this paper and Gaunt et al., 2003). A null mutation in a single cdx gene will therefore cause only a limited posterior shift in the total cdx protein concentration gradient and hence may produce only limited posterior shifts in Hox expression boundaries. These shifts may, however, be enhanced when multiple cdx genes are mutated, as has been reported (van den Akker et al., 2002).
Fig. 6. Possible scheme for the establishment of Hox expression
boundaries along instructional cdx gradients. During gastrulation, the
cdx2 and cdx4 gradients regress posteriorly along the developing axis
(indicated here by a vertical arrow). At early times only 3'-located Hox genes are expressed and their boundaries may become differentially sorted by the cdx protein gradients as they then exist over prospective anterior parts of the embryo. This may occur, for example, because each Hox gene is
activated by the products of more 3' Hox genes (Hooiveld et al., 1999;
McNulty et al., 2005), but only above a threshold cdx protein concentration.At later times, more 5'-located Hox genes become expressed and their boundaries become sorted over the more posterior structures which are then displaying the cdx gradients. The protein gradients from all three cdx genes likely overlap (this paper and Gaunt et al., 2003). A null mutation in a single cdx gene will therefore cause only a limited posterior shift in the total cdx protein concentration gradient and hence may produce only limited posterior shifts in Hox expression boundaries. These shifts may, however, be enhanced when multiple cdx genes are mutated, as has been reported (van den Akker et al., 2002).
Члены кластера Hoxc экспрессируются в разных пространственных доменах вдоль длины нервной трубки и позиции их чувствительны к концентрациям Fgf (Liu et al., 2001). Т.к. наиболее высокие урвони Fgf обнаруживаются только в хвостовой области (Dubrulle and Pourquiй, 2004), то возможно. что роль Fgf в формировании паттерна обычно опосредуется через градиенты распада cdx. И cdx1 и cdx4 чувствительны к Fgf (Bel-Vialar et al., 2002).
В коротких и промежуточных зародышевых дисках насекомых, также как и у позвоночных, ген caudal сходным образом экспрессируется в задней ростовой зоне, в то время как происходит сегментация (Dearden and
Akam, 2001; Shinmyo et al., 2004). Остается неясным, однако, формируются ли градиенты за счёт распада беспозвоночных хвостовых белков, и могут ли они создавать инструктивные градиенты для установления границ экспрессии генов.
Сайт создан в системе
uCoz DEBORAH DRAGE and RICHARD C. TRUBSHAW
DEBORAH DRAGE and RICHARD C. TRUBSHAW