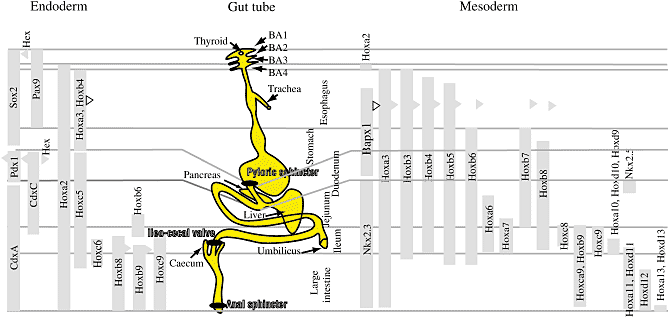
Интересное наблюдение относительно сигналов, меняющихся с помощью энтодермы и мезодермы (see below), фактически говорит о том, что энтодерма скользит кзади по мезодерме (Рис. 3). Напр., N. Le Douarin показала, что частички угля оказываются во всех трех зародышевых слоях на уровне конца сомита 2, на уровне шеи в нервной трубке и на уровне гортани в мезодерме и еще более кзади в brunchi в энтодерме (Le Douarin, 1964; Tremblay and Zaret, 2005). Это общее скольжение кзади энтодермы относительно мезодермы и мезодермы относительно нейроэктодермы наиболее выражено в задней и боковых частях (Catala et al., 1996, Le Douarin, 1964a). Соответственно, на молекулярном уровне передние границы Hox генов являются наиболее ростральными в нейроэктодерме, затем сомитах, затем в латеральной пластинке и затем в энтодерме (Fig. 3) (Burke et al., 1995). Сдвиг в том же самом направлении описан между эктодермой и висцеральной мезодермой у
(Tremml and Bienz, 1989). Помимо этих движений, Smith and Tabin (Smith and Tabin, 2000) показали, что мезодермальные клетки мигрируют индивидуально перед стадией 13 сомитов и могут таким образом клонально вносить вклад в разные органы кишечника. После этой стадии, хотя клоны и могут занимать разные радиальные слои, они ограничиваются одним органом.
Культивируемая сама по себе энтодерма выживает очень плохо (Le Douarin and Bussonnet, 1966, Okada, 1953, Okada, 1954a, Okada,
1954b, Sumiya and Mizuno, 1974, Takata, 1960) или поддерживается в течение очень короткого времени (Kumar et al., 2003, Wells and Melton, 2000)
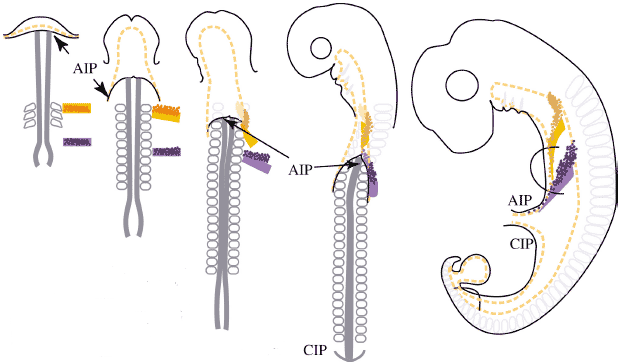 Fig. 3. Morphogenetic movements during gut tube closure in chick. Schematized ventral views of 3-somite, 10-somite, 20-somite and 28-somite
embryos as well as an embryo which has completed gut tube closure at the end of somitogenesis. The anterior intestinal portal (AIP)
forms at the beginning of somitogenesis and the
caudal intestinal portal (CIP) at about 28-somite stage. Their progression
towards the middle of the embryo closes the gut tube delineated by
yellow lines. The endoderm at the level of somite 1 is marked by light
orange fill. Dark orange dots represent the lateral plate mesoderm from the same level. Somite 6 level is similarly labelled: light purple fill for endoderm and dark purple dots for mesoderm. The enhanced posterior shift of more lateral tissues is seen for endoderm and mesoderm as well as the enhanced posterior shift of endoderm relative to somites and mesoderm. As a result endoderm from a given A-P location contributes to different organs. The same is true for mesoderm. Moreover, endoderm and mesoderm of the same A-P origin end-up in different positions.
Fig. 3. Morphogenetic movements during gut tube closure in chick. Schematized ventral views of 3-somite, 10-somite, 20-somite and 28-somite
embryos as well as an embryo which has completed gut tube closure at the end of somitogenesis. The anterior intestinal portal (AIP)
forms at the beginning of somitogenesis and the
caudal intestinal portal (CIP) at about 28-somite stage. Their progression
towards the middle of the embryo closes the gut tube delineated by
yellow lines. The endoderm at the level of somite 1 is marked by light
orange fill. Dark orange dots represent the lateral plate mesoderm from the same level. Somite 6 level is similarly labelled: light purple fill for endoderm and dark purple dots for mesoderm. The enhanced posterior shift of more lateral tissues is seen for endoderm and mesoderm as well as the enhanced posterior shift of endoderm relative to somites and mesoderm. As a result endoderm from a given A-P location contributes to different organs. The same is true for mesoderm. Moreover, endoderm and mesoderm of the same A-P origin end-up in different positions.
а позднее сообщалось об обнаружении в таких культурах немногих мезенхимных клеток (Gualdi et al., 1996) (Rossi et al., 2001).
С момента, как Sumiya and Mizuno (1974) сообщили о культуре, завернутой в вителлиновую мембрану, они стали культивироваться в присутствии мезенхимы или трансплантатов в хорионалантоис (Butler, 1935, Le Douarin, 1964a). Гетерологичная спланхническая мезенхима в общем позволяет выживать и даже собственно дифференцировку, но мезодерма из зачатков конечностей, сомитная или цефалическая плохо ей замещают (Le Douarin, 1964a, Le Douarin and Bussonnet, 1966, Le Douarin and Wolff, 1967). Молекулярная природа некоторых из сигналов, продуцируемых локально спланхнической мезодермой, недавно установлена. Ранние эксперименты Nicole Le Douarin (Le Douarin,
1964a, Le Douarin, 1964b, Le Douarin, 1964c)показали, что сигналы от мезодермы сердца необходимы для индукции развития печени. Презумптивная территория печени, AIP, трансплантированные на др. мезодермальные территории, могут развиваться автономно после стадии 6 сомитов, но нуждаются в присутствии кардиальной мезодермы перед этой стадией. Т.к. кардиальная мезодерма не индуцирует печень при комбинировании её с др. областями энтодермы, то эти сигналы являются скорее пермиссивными, чем инструктивными (Le Douarin, 1975).
Эти свойства недавно были подтверждены на мышах (Gualdi et
al., 1996). Очевидно, что у мышей в отсутствии кардиальной мезодермы, энтодерма из AIP экспрессирует панкреатические маркеры (Deutsch et al., 2001). FGF1 и FGF2, которые экспрессируются в кардиальной мезодерме, м. индуцировать печеночные маркеры в AIP, а растворимые формы FGF рецепторов, которые оказывают доминантно негативные эффекты, блокируют индукцию печени за счет кардиальной мезодермы. Недавно было предположено, что FGFs, посылаемые кардиальной мезодермой, также необходимы и для развития лёгких (Serls et al., 2005) и что они могут играть инструктивную роль, т.к. разные концентрации индуцируют разные органные маркеры. Поперечная перегородка (septum transversum), еще одно производное lateral
plate mesoderm (LPM), участвует в индукции печени (Rossi et al., 2001) и в более позднем её поддержании (Le Douarin, 1963, Le Douarin, 1964a, Le Douarin, 1964c). Она продуцирует BMPs. Воздействие noggin, ингибитором BMP, блокирует конвергентную способность кардиальной мезодермы и поперечной перегородки в индукции печеночных маркеров. На более поздних стадиях развития, производные LPM сигналы, как известно, существенны для поддержания экспрессии Pdx1 в энтодерме поджелудочной железы. Мыши, мутантные по Isl1 (Ahlgren et al., 1997)и N-cadherin (Esni et al., 2001; Edsbagge et al., 2005), двум генам, экспрессирующимися в LPM, не обладают LPM конвергенцией вокруг дорсальной панкреас и обнаруживают резкое подавление экспрессии Pdx1, а также арест развития органа после инициального отпочкования зачатка. FGF10, ген, экспрессируемый в мезенхиме вокруг панкреатического зачатка от момента инициации отпочкования зачатка и далее, необходим для поддержания экспрессии Pdx1 (Bhushan et al., 2001).
В дополнение к LPM, ещё два мезодермальных компонента контактируют с энтодермой, обеспечивая пермиссивными сигналами. Хорда, как было показано, необходима и достаточна для индукции панкреатических эндокринных и экзокринных маркеров на стадии 10 сомитов в презумптивной энтодерме панкреас (Kim et al., 1997). Она посылает пермиссивные сигналы, т.к. она не может индуцировать панкреатические маркеры в более задней энтодерме. FGF2 и activin, как было показано, воспроизводят сигналы хорды, но пока неищвестно, действуют ли они и
in vivo (Hebrok et al., 1998). Хотя ActRIIB
-/- и ActRIIA
+/-B
-/- мутантны и обнаруживают трансформации в более передние части задних частей желудка, но неясно, обусловлен ли этот эффект восприятием сигналов от хорды (Kim, 2000). В самом деле, экзокринная часть поджелудочной железы не затрагивается у этих мутантов, хотя хорда контролирует также и индукцию экзокринной части панкреас. Кроме того, LPM вокруг поджелудочной железы модифицируется у этих мутантов. Неизвестно, может ли хорда также посылать пермиссивные сигналы в др. регионы кишки и выполнять общую роль в формировании дорсо-вентрального паттерна энтодермы. Эндотелий кровеносных сосудов также необходим для собственно дифференцировки дорсальных частей панкреас и печени (Lammert et al., 2001, Matsumoto et al., 2001). В случае печени, инициальная спецификация происходит нормально в отсутствие эндотелиальных клеток, но клетки печени не способны пролиферировать и проникать в мезенхиму поперечной перегородки (Matsumoto et al., 2001). Так, для панкреас аорта необходима для дифференцировки эндокринных клеток и морфогенеза дорсального панкреатического зачатка (Lammert
et al., 2001; Yoshitomi and Zaret, 2004). Недавние доказательства подтверждают, что вителлиновые вены, которые находятся в тесной близи к вентральному зачатку поджелудочной железы, не нужны для инициации вентрального зачатка (Yoshitomi and
Zaret, 2004). Возможно, что эта ассоциация также важна и для других пищеварительных органов, которые также обладают ранней ассоциацией с эндотелиальными клетками (Matsumoto et al., 2001). Пока неясно, любые ли эндотелиальные клетки обладают подобной индуктивной способностью.
Lateral plate mesoderm located at different A-P
positions sends different instructive information to
endoderm
У
Drosophila, паттерн энтодермы (средняя кишка) формируется благодаря регионализованным сигналам из мезодермы (Bienz, 1994,
Bienz, 1997). У позвоночных имеет место длительное общение между энтодермой и мезодермой, что может быть проиллюстрировано на примере печени и сердца. Уже давно отмечено, что эти органы развиваются вместе (Hunt, 1932, Willier and Rawles, 1931). Вскоре после гаструляции передняя энтодерма, но не задняя энтодерма посылает сигналы, которые индуцируют кардиомиоциты в мезодермальных клетках, которые покидают первичную полоску (Schultheiss et al., 1997, Schultheiss et al., 1995). Wnt-ингибиторы, экспрессирующиеся в передней энтодерме блокируют сердце-репрессирующую активность задних Wnts (Marvin et al., 2001, Tzahor and Lassar, 2001; Foley and Mercola, 2005; Schneider and Mercola, 2001).
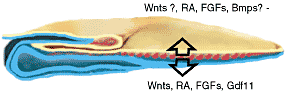 Fig. 4. The mesoderm coordinates A-P patterning in the three germ
layers. A-P patterning of endoderm and neurectoderm is under the
control of mesodermal signals. RA and FGFs pattern the two germ layers.
Although Wnts play a role in A-P patterning of the neural tube questions
remain about the time of action and about its role in endoderm. TGF в
family members are involved in patterning both layers but the mechanism
and precise members involved need to be better characterized.
Fig. 4. The mesoderm coordinates A-P patterning in the three germ
layers. A-P patterning of endoderm and neurectoderm is under the
control of mesodermal signals. RA and FGFs pattern the two germ layers.
Although Wnts play a role in A-P patterning of the neural tube questions
remain about the time of action and about its role in endoderm. TGF в
family members are involved in patterning both layers but the mechanism
and precise members involved need to be better characterized.
На ранних сомитных стадиях сердце передает сигналы обратно в энтодерму, чтобы индуцировать гепатоциты. Позднее septum transversum оказывается необходимой для роста печени. Этот и др. примеры ведут к заключению, что позиционная информация в ЖКТ не содержится ни в энтодерме, ни в мезодерме, обе они обладают частичной информацией, которая варьирует в зависимости от времени (Yasugi, 1993).
Всё ещё неясно, имеется klb общая схема формирования паттерна энтодермы, которая координирует со всей A-P осью. Показано, что на сомитных стадиях, когда большинство позиционных маркеров индуцируется в энтодерме, LPM посылает инструктивные сигналы в энтодермальный слой, по крайней мере от двенадцатиперстной кишки до задней части тонкого кишечника (Kumar et al., 2003). В отсутствие LPM, энтодерма сохраняет нативное энтодермальное состояние, но не включает и не сохраняет позиционных маркеров (Horb and Slack, 2001, Kumar et al., 2003). Она сохраняет свою исходную A-P качественную особенность при ассоциации с более передней энтодермой, но она формирует новый паттерн, когда ассоциирует с более задней LPM. Это очень сходно с механизмом формирования паттерна нервной трубки на той же стадии, т.к. было показано, что сомиты и Гензеновский узелок могут индуцировать более задние, но не более передние судьбы (Grapin-Botton et al., 1997, Itasaki et al., 1996, Liu et al., 2001, Muhr et al., 1997). Сомиты обладают общей с LPM способностью индуцировать задние маркеры в энтодерме (Kumar et al., 2003). Это указывает на то, что мезодерма координирует формирование паттерна всех трёх зародышевых листков (Рис. 4). Т.к. в нервную трубку Гензеновский узелок также посылает posteriorizing сигналы на сомитных стадиях, то было бы интересно посмотреть, не играет ли узелок такую же роль позднее в формировании паттерна энтодермы (Liu et al., 2001). Пластичность на эквивалентных стадиях наблюдалась у лягушек, у которых на стадии 22-25, но не на стадии 28 энтодерма теряет экспрессию Xlhbox8/Pdx1, если ассоциирована с задней мезодермой (Zeynali et al., 2000). Градиент передаваемых сигналов вдоль A-P оси сравним с наблюдениями Nicole Le Douarin, которая трансплантировала энтодермальный зачаток печени после стадии 6 сомитов в спланхноплевру и показала, что более каудальные трансплантаты дают менее развитые эпителиальные печеночные тяжи (Le Douarin, 1963, Le Douarin, 1964c). Это согласуется также с наблюдением, что энтодерма тонкого кишечника теряет очень рано свою способность формировать заново паттерн, когда ассоциирует с мезенхимой из др. регионов (Andrew and
Rawdon, 1990, Gumpel-Pinot et al., 1978, Yasugi, 1993, Yasugi
and Mizuno, 1978, Yasugi et al., 1991). Всё ещё неясно, действительно ли верно наблюдаемое posterior dominance мезодермы вдоль всей A-P оси, т.к. только незначительные отклонения обнаружены в литературе. Дно глотки, трансплантированное в спланхноплевру той же стадии и позднее, обнаруживается в стенке тонкого кишечника или в стенке тела, формирует при этом щитовидную железу, тимус, мышечную и железистую части желудка, печень, экзокринную поджелудочную железу и кишечник (Le
Douarin and Bussonnet, 1966, Le Douarin and Wolff, 1967). Передний маркер Sox2 (Рис. 2) индуцируется в тонком кишечнике вплоть до ст. E4
(соответствет 11.5 dpc у мышей) (Ishii et al., 1998).
Пластичность, по-видимому, теряется задолго до рождения Ishii с сотр. показали, что эпителий желудка posteriorized с помощью эпителия тонкого кишечника вплоть до E4, но на ст. E6 (примерно 12.5dpc), хотя сигналы всё ещё продуцируются мезенхимой, энтодерма теряет свою способность экспрессировать кишечные маркеры CdxA и sucrase (Ishii et al., 1997, Ishii et al., 1998) (Haffen et al., 1982, Ishizuya-Oka and Mizuno, 1984). Соотв., 14.5dpc эпителий желудка или легких, по-видимому, не испытывают влияния со стороны мезенхимы тонкого кишечника (Duluc et al., 1994). Мезенхимные сигналы, по-видимому, персистируют (perdure) после потери пластичности (Ishii et al., 1997). Не всегда ясно, относятся ли идентифицированные сигналы к формированию общего A-P паттерна или к локальным событиям. Напр., железы желудка могут развиваться во многих эпителиях, включая заднюю часть тонкого кишечника вплоть до поздних стадий (E9). В этом случае железистые структуры формируются, но не экспрессируют pepsinogen, но сохраняют sucrase (Takiguchi-Hayashi and Yasugi,
1990, Yasugi et al., 1985). Информация по генерации структуры находится в мезенхиме, но A-P информация может находиться в эпителии. В таком случае, мышечная мезенхима желудка, смешанная в 1:9 с железистой мезенхимой желудка, блокирует способность индуцировать железы, даже если мышечные клетки желудка находятся в отдалении от эпителия, подтверждая тем самым, что локальные репрессивные секретируемые сигналы действуют на расстоянии (Urase and Yasugi, 1993).
Качественные A-P особенности параксиальной мезодермы связаны с сегментацией и предпопределяются, когда формируются сомиты (Dubrulle et al., 2001, Kieny et al., 1972, Nowicki and Burke, 2000, Zakany et al., 2001). Имеется, по-видимому, более поздняя пластичность в отношении качественных особенностей A-P в соматоплевре, т.к. когда индуцируется дополнительная конечность путем воздействия FGF-нагруженных кусочков, между крылом и конечностью, то экспрессия Hox генов в соматоплевре или сдвинута кзади, так что напоминает паттерн крыла, или сдвинута кпереди, так что напоминает паттерн конечности (Cohn et al., 1997). Более того LPM среда изменяет качественные A-P особенности в изолированных сомитных клетках, которые мигрируют в LPM, чтобы сформировать мышцы конечностей (Nowicki and Burke, 2000). Ничего подобного неизвестно о спланхноплевре.
Molecular basis of endoderm A-P patterning
Ретиноевая кислота индуцирует задние характеристики энтодермы, как и в нейроэетодерме. В нервной трубке FGFs (Cox and Hemmati-Brivanlou, 1995;
Kengaku and Okamoto, 1995; Lamb and Harland, 1995; Mathis,
et al., 2001; Storey, et al., 1998), Wnts (Erter, et al., 2001;
Mcgrew, et al., 1997; Nordstrom, et al., 2002) и retinoic acid
(RA) (Maden, 1999),как известно, индуцируют задние судьбы градированным способом (Bel-Vialar, et al., 2002; Liu, et al., 2001). Из TGFβ ctvtqcndf Growth/Differentiation Factor GDF11, как полагают, ответственен за индукцию наиболее задних судеб в сотрудничестве с FGFs (Liu, et al., 2001; Mcpherron, et al., 1999).
Так, в нервной трубке RA оказывает posteriorizing эффекты на энтодерму. На ст. гаструляции у эмбрионов кур она способна репрессировать передний маркер Otx2 (Bally-Cuif, et al., 1995). Роль RA
в формировании энтодермальных органов изучалась на рыбках данио, Xenopus, мышах и курах (Chen, et al., 2004; Kumar, et al.,
2003; Molotkov, et al., 2005; Stafford, et al., 2004; Stafford and
Prince, 2002). Эти работы сконцентрированы в основном на поджелудочной железе и было показано, что ретиноевая кислота важна на ст. гаструляции, чтобы индуцировать этот орган и контролировать баланс между эндокринными и экзокринными клетками. Хотя и проанализированные менее тщательно, но две из этих публикаций указывают на то, что RA контролирует положение энтодермальных органов вдоль A-P оси, как это имеет место в нервной трубке. У рыбок данио RA индуцирует передний сдвиг в экспрессии энтодермальныхы генов по всей передней кишке (down to duodenum/pancreas) и блокирует путь, предупреждающий индукцию более задних судеб (Stafford
and Prince, 2002). У кур, Kumar et al. (Kumar, et al., 2003)
наблюдалось, что RA индуцирует Pdx1, pancreato-duodenal маркер в stomach/esophagus, а также расширяет экспрессию CdxA, маркера тонкого и толстого кишечника, вплоть до пищевода. Однако, эффекты, описанные у кур, появляются на значительно более поздних стадиях, когда сомиты уже сформированы. Хотя потребность в RA для формирования паттерна энтодермы не была предметом рассмотрения данной работы, очевидно, что энтодерма всё ещё реагирует на RA на стадии 10 сомитов в отличие от рыб и Xenopus. Многие A-P маркеры энтодермы индуцируются на этой стадии. Она также контролирует формирование A-P паттерна энтодермы у Amphioxus (Escriva, et al., 2002; Schubert, et al., 2005), но эти исследования противоречат предыдущим находкам, показавшим, что у Xenopus RA не влияет на экспрессию панкреатического маркера Pdx1 после воздействия на эмбриона целиком на ст. 12 и 25 (Zeynali and Dixon, 1998). Более позднее действие RA у эмбрионов кур согласуется с задне-передним градиентом RA, присутствие которого предположено в LPM эмбрионов кур на ст. 10 сомитов (Swindell, et al., 1999). В самом деле,
Raldh2, энзим, ответственный за синтез RA synthesis, экспрессируется в полоске и позднее в сомитах и LPM туловища, тканях, которые обладают posteriorizing активностью. RA , т.о., синтезируется только в туловище.
Cyp26, энзим, ответственный за деградацию RA, часто экспрессируется в тканях, которые реагируют на RA, находится в латеральной энтодерме. Его экспрессия формирует градиент на ст. 14, с высоким уровнем экспрессии в первом сомите и постепенным снижением в направлении сомита 15. Количество сигнала, воспринимаемого боле5е каудально мб
более высокое из-за уменьшения скорости деградации (Blentic, et al., 2003). Пока неясно, формирует ли RA паттерны всей энтодермы вдоль оси. В нервной трубке RA играет роль только в регуляции генов от заднего мозга (уровень глотки) до уровня шеи (Hoxc-6). Она подавляет более задние гены Hox
in vivo и in vitro (Bel-Vialar, et al., 2002; Liu, et al., 2001; Simeone, et al., 1991). В энтодерме, RA, по-видимому, индуцирует каудальный гомолог
CdxA у кур и не влияет у рыбок данио (Kumar, et al., 2003; Stafford and Prince, 2002). В каждом из этих случаев неясно, действует ли RA непосредственно на энтодерму или косвенно через мезодерму. Лучшее доказательство прямого действия RA на эктодерму получено при исследовании Hoxb1 промотора, который регулируется с помощью RA в энтодерме и эктодерме посредством двух разных промоторных элементов (Huang, et al., 2002; Huang, et al., 1998). Такие элементы не были идентифицированы в промоторах
Pdx1 или
CdxA/1. Этот путь может взаимодействовать с FGF путем в энтодерме во время гаструляции, т.к. путь FGF регулирует экспрессию RA рецепторов и энзимов, которые контролируют синтез и деградацию RA (Shiotsugu, et al., 2004) и напротив передача сигналов RA регулирует экспрессию FGF рецепторов.
FGFs and Wnts, posterior inducers in the nervous
system may act on endoderm patterning
В нервной системе на стадии сомитов RA ответственна за формирование паттерна на уровне заднего мозга и на уровне шеи. Более каудально экспрессия Hox генов индуцируется с помощью FGFs
(в основном 2 и 4 и менее эффективно 8). которые действуют частично посредстом Cdxs (Bel-Vialar et al., 2002; Charite et al., 1998; Liu, et al., 2001). В энтодерме мышей, FGF4 (но неt FGF2, 5 or 8)
действует в передней энтодерме в поздней гаструле (7.5 dpc), индуцируя маркеры, которые характерны для задней половины эмбриона. FGF4 экспрессируется в задней части первичной полоски и позднее в хвостовой почке (Niswander and Martin, 1992). Эмбрионы, нокаутные по Fgf4, останавливаются в развитии перед гаструлой (Feldman et al., 1995) и следовательно, потребность в FGF4 для формирования A-P паттерна не может быть протестирована. Тогда как в нервной системе FGF2 и 4 наиболее мощные задние индукторы. Наши недавние эксперименты подтвердили, что FGFs функционируют также позднее в формировании паттерна средней кишки (Dessimoz et al., unpublished). FGFs могут оказывать пермиссивное действие и поддерживать клетки в состоянии чувствительности к сигналам (Mathis et al., 2001; Nordstrom
et al., 2002). FGF response elements, которые связывают ets транскрипционные факторы (нижестоящие мишени FGFs), недавно были выявлены в Xcad3 (CdxB) промоторе у Xenopus. CdxB может т.о., быть непосредственной мишенью в энтодерме для передачи сигналов FGF (Haremaki et al., 2003). Pdx1 также может быть непосредственной мишенью для сигналов FGF4.
Wnts, в присутствии FGFs, играют роль в формировании A-P паттерна нервной системы (Holland, 2002; Mcgrew et al., 1997; Nordstrom
et al., 2002).
Wnt8 экспрессируется в каудальной параксиальной мезодерме на ст. гаструлы и головной складки и активирует задние маркеры градированным способом в изолированной нервной трубке. Неясно, поддерживает ли он передачу сигналов на более поздних стадиях.
Wnt3A экспрессируется в хвостовой почке, также как
wnt8 у ряда позвоночных (Holland, 2002).
Cdx1/A является непосредственной мишенью канонической передачи сигналов Wnt, которая важна для индукции этого гена в первичной полоске, сомитах (wnt3A) и возможно энтодерме (Ikeya and Takada, 2001; Lickert et al., 2000;
Lickert and Kemler 2002). Возможно существует петля обратной связи, т.к.
CdxB, каудальный ген у кур, м. индуцировать
Wnt8c, когда неправильно экспрессируется в сердце (Ehrman and Yutzey, 2001). Т.к.
Cdx1 индуцируется позднее во время развития в энтодерме (14 dpc) то было бы интересно знать, зависит ли индукция ранних энтодермальных маркеров также от передачи сигналов Wnt. Ни один из изолированных промоторных элементов
Cdx1 , включая и те для сайтов мишеней пути Wnt, не управляют энтодермальной экспрессией (Lickert and Kemler, 2002).
Интересно, что у
C. elegans, все, кроме одного деления в E
клоне, происходят вдоль A-P оси. Эти деления асимметричны, так что только передняя клетка наследует Tcf-родственный белок POP-1 (Lin et al., 1998). В первом асимметричном делении дробления EMS, которое генерирует энтодермальный предшественник E posteriorly, это обусловлено продукцией MOM-2 Wnt лиганда P2 задней клеткой (Lin et al., 1998). Более поздние деления нуждаются в MOM-5/Frizzled, но не в MOM-2/Wnt (Park and Priess, 2003). У
Drosophila, Wingless секретируется мезодермой, окружающей энтодерму, где он индуцирует
labial, а позднее copper клетки при низких уровнях, тогда как ближе к месту продукции он репрессирует
labial и индуцирует дифференцировку крупных плоских клеток (Hoppler and Bienz, 1995).
A role for BMPs in endoderm A-P patterning: differences
between endoderm and neurectoderm?
У Drosophila, Hox гены регионально экспрессируются в мезодерме и контролируют локальную секрецию Decapentaplegic
(Dpp). Dpp экспрессируется в специфической полоске в мезодерме средней кишки, по соседству с Wg. Он регулирует экспрессию Hox генов в мезодерме и энтодерме (Immergluck et al., 1990, Staehling-Hampton and Hoffmann, 1994, Staehling-Hampton et al., 1994, Tremml and
Bienz, 1989)(Tremml and Bienz, 1989; Immergluck et al.,1990;
(Panganiban et al., 1990). Экспрессия labial в энтодерме зависит от кооперации Hox связывающих сайтов, которые связывают отвечающие элементы в labial и DPP (Marty et al., 2001). LEF1 и DPP/mad связывающие сайты в промоторе ultrabithorax непосредственно контролируют этот ген (Riese et al., 1997).
У позвоночных было показано, что BMP2, 4 и 7 , а также activin
расширяют экспрессию Pdx1 и CdxA в энтодерме кпереди (Kumar et al., 2003). Кроме того, noggin и follistatin блокируют индукцию задних генов, выявляемых с помощью LPM, подтверждая, что вовлечен BMP скорее, чем activin (activin не блокируется с помощью noggin). Как уже отмечалось, во время морфогенеза кишечника более латеральная энтодерма в конечном итоге вносит вклад в более задние ткани, вполне возможно, что эффект BMPs проявляется в латеральной трансформации. У рыбок данио, Tiso et al. (Tiso et al., 2002) продемонстрировали, что swirl (BMP2b) мутанты обнаруживают общую редукцию качественных особенностей в задней части энтодермы. Напротив, мутанты chordino увеличенную заднюю кишку и уменьшенную переднюю часть ЖКТ. Хотя эффект BMP2b, по-видимому, является очень ранним у рыб, т.к. экспрессия гена hairy her5 в передней энтодерме затронута у мутантов swirl и chordino в 80% epiboly (late gastrulation), но было установлено, что activin не posteriorize 7.5 dpc энтодерму мышей (gastrulation). Должны существовать различия во времени формирования A-P паттерна у этих двух видов, как это наблюдалось в случае ретиноевой кислоты (Wells and Melton, 2000). Две первые публикации показали, что эти лиганды способны индуцировать экспрессию генов, характерных для множественных регионов вдоль A-P оси и поэтому они скорее всего действуют в виде градиентов, хотя это еще не доказано. Можно ожидать, увидеть градированную экспрессию одного или нескольких BMPs или activin вдоль A-P оси на изученных здесь стадиях. В самом деле, LPM, соседствующая с пресомитной мезодермой, строго экспрессирует BMP4 причем экспрессия прогрессивно ослабляется в LPM, соседствующей с полностью сегментированными сомитами (Pourquie et al., 1996, Reshef et al., 1998, Schultheiss and Lassar, 1997) (our unpublished results). Кроме того, экспрессия множественных BMPs выявляется в мезенхиме, окружающей энтодерму (Roberts et al., 1995, Smith et al., 2000, Solloway and Robertson, 1999, Winnier et al., 1995), тогда как BMP рецепторы BMPRIA и BMPRII экспрессируются в энтодерме (Mishina et al., 1995, Roelen et al., 1997). Инактивация activin receptor II у мышей выявляет потребность в этих рецепторах для формирования A-P паттерна передней кишки (Kim et al., 2000). Т.к. Gdf11 играет роль в индукции задних генов в нервной пластинке, то было бы интересно посмотреть, вовлекается ли GDF в posteriorization энтодермы (Liu et
al., 2001, McPherron et al., 1999).
Roberts et al. (Roberts et al., 1995) показали, что
Shh индуцирует Hox гены в задней части задней кишки.
Shh первоначально экспрессируется в энтодерме CIP, когда кишка сворачивается и позднее его экспрессия распространяется на большую чать кишечной трубки. Преждевременная активация
Shh в кишечной энтодерме индуцирует сдвиг кпереди экспрессии
Hoxd11 и Hoxd13 в мезодерме. Этот сдвиг ограничен , т.к. области впереди желточного стебелька не компетентны экспрессировать эти гены на этой стадии. Неясно, как действие Shh ограничивается, т.к. этот ген экспрессируется в AIP, где он не индуцирует задних Hox генов. Т.к. Shh индуцирует также BMPs в мезодерме, то способность Shh индуцировать задние гены может быть связана с BMPs.
Conclusion
Beyond characterizing the interactions between the retinoic
acid, Wnt, FGF and BMP pathways in endoderm patterning, new
directions of research may go towards further characterization of
endoderm-mesoderm interactions in light of the sliding movements
observed by Nicole Le Douarin. Fate mapping the AIP with modern
techniques should in particular solve the question as to whether
single cells of the AIP can contribute to different organs.
The control of A-P identity in endodermal cells is important not
only during development but must be maintained in the adult. The loss of identity leads to metaplasia which often evolve in neoplasia.
Mice which have lost one copy of Cdx2 in their genome develop
adenomatous polyps, mostly in the proximal colon (Beck et al.,
2003, Beck et al., 1999, Chawengsaksophak et al., 1997). These
polyps contain areas were Cdx2 is not expressed although there
is no loss of heterozygosity. These Cdx2 -negative domains show
a continuous sequence of all digestive tract epithelia from colon to
esophagus suggesting a gradient of signaling. Dysplasia is observed
in 20% of lesions. Although Cdx2 expression is often lost in colon
cancer, it is rarely mutated (da Costa et al., 1999, Mallo et al., 1997,
Yagi et al., 1999). Another example of induced metaplasia mimics
a human affection. When Cdx2 is ectopically activated in the
stomach, intestinal metaplasia is induced (Silberg et al., 2002).
Cdx2 expression is observed in most gastric intestinal metaplasia
and gastric carcinoma (Almeida et al., 2003, Bai et al., 2002, Seno
et al., 2002). In the esophagus, 10% of Barrett metaplasia, which
consist in gastric tissue in the esophagus, evolve in esophagal
cancer (OMIM 109350). The role of Cdx up- or down-regulation in
the evolution to neoplasia and the factors that cause it are interesting
questions for the future. In particular, since the mesenchyme
controls Cdx expression during development, the role of the
stroma would be worth investigating.
Сайт создан в системе
uCoz
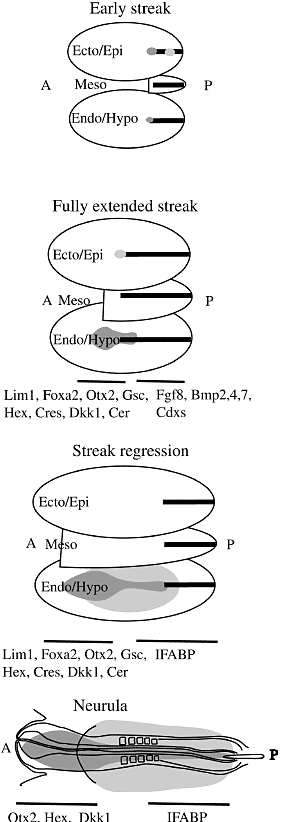 Fig. 1. Spatial organization of endoderm at gastrulation and headfold stage in the chick. The three layers are schematically dissociated, epiblast/ectoderm on top, mesoderm in the middle, hypoblast/endoderm at the bottom. Anterior/midline endoderm is in dark grey and posterior/lateral endoderm is in pale grey. The presence of endodermal cells in transit in the middle layer has not been assessed. The dark line is the primitive streak. A refers to anterior and P to posterior. Anterior markers are the transcripts for the secreted proteins Cerberus 1 (Cer) (Belo et al., 1997; Biben et al., 1998; Chapman et al., 2002), Dickkopf1 (Foley et al., 1997; Glinka et al., 1998; Pearce et al., 1999), Crescent (Pfeffer et al., 1997) and the homeobox transcription factors Orthodenticle homeobox (Otx) 2 (Ang et al., 1994; Bally-Cuif et al., 1995), Goosecoid (Izpisua-Belmonte et al., 1993; Blum et al., 1992), Hex (Thomas et al., 1998; Yatskievych et al., 1999) and Lim Homeobox 1 (Lim1) (Chapman et al., 2002; Shawlot and Behringer, 1995) as well as FoxA2 (Alexander and Stainier, 1999; Ang et al., 1993; Ruiz i Altaba et al., 1995), Her5 (Bally-Cuif et al., 2000). Many of these genes are also expressed in the primitive endoderm in the AVE in addition to definitive endoderm (Rodriguez et al., 2001). Posterior markers of endoderm are more scarce but one of them is Intestinal fatty acid binding protein transcript (IFABP) (Wells and Melton, 2000). Many posterior markers like Fgf8, Bmp 2,4,7, Cdxs (Chapman et al., 2002; Marom et al., 1997) are expressed in the posterior streak where layers can not be distinguished
Fig. 1. Spatial organization of endoderm at gastrulation and headfold stage in the chick. The three layers are schematically dissociated, epiblast/ectoderm on top, mesoderm in the middle, hypoblast/endoderm at the bottom. Anterior/midline endoderm is in dark grey and posterior/lateral endoderm is in pale grey. The presence of endodermal cells in transit in the middle layer has not been assessed. The dark line is the primitive streak. A refers to anterior and P to posterior. Anterior markers are the transcripts for the secreted proteins Cerberus 1 (Cer) (Belo et al., 1997; Biben et al., 1998; Chapman et al., 2002), Dickkopf1 (Foley et al., 1997; Glinka et al., 1998; Pearce et al., 1999), Crescent (Pfeffer et al., 1997) and the homeobox transcription factors Orthodenticle homeobox (Otx) 2 (Ang et al., 1994; Bally-Cuif et al., 1995), Goosecoid (Izpisua-Belmonte et al., 1993; Blum et al., 1992), Hex (Thomas et al., 1998; Yatskievych et al., 1999) and Lim Homeobox 1 (Lim1) (Chapman et al., 2002; Shawlot and Behringer, 1995) as well as FoxA2 (Alexander and Stainier, 1999; Ang et al., 1993; Ruiz i Altaba et al., 1995), Her5 (Bally-Cuif et al., 2000). Many of these genes are also expressed in the primitive endoderm in the AVE in addition to definitive endoderm (Rodriguez et al., 2001). Posterior markers of endoderm are more scarce but one of them is Intestinal fatty acid binding protein transcript (IFABP) (Wells and Melton, 2000). Many posterior markers like Fgf8, Bmp 2,4,7, Cdxs (Chapman et al., 2002; Marom et al., 1997) are expressed in the posterior streak where layers can not be distinguished