OSNs обладают строгой и внутренне присущей способностью находить один другого и соединяться без образования синапсов со своими обычными партнерами. Роль для обоняния окружающих клеток или глии нельзя исключить в этих экспериментах.
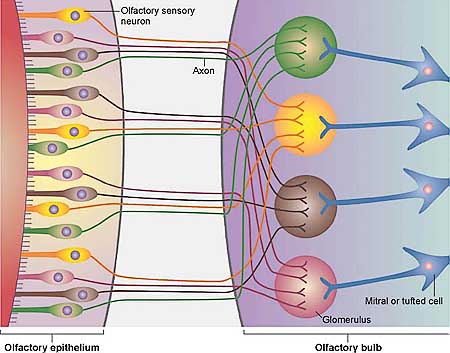
Обонятельный эпителий постоянно регенерирует самого себя в течение всей жизни. Часто полагают, что OSNs продуцируются из клеток предшественников автоматически и регулярно, подобно эпидермису или эпителию ЖКТ или респираторного тракта. То, что OSNs имеют продолжительность жизни в 30 или 90 дней, пропитана литература, но эти количества приводятся в результате поверхностной интерпретации данных (таких как Graziadei et al. 1979). 90-дневная продолжительность жизни, которая иногда даже присутствует как период полужизни, была предложена Mackay-Sim & Kittel (1991a,b), к их собственному разочарованию (Mackay-Sim 2003). 90-дневный промежуток времени позднее исследовался этими авторами и привел к более низким значениям продолжительности жизни; определенно это не период полу-жизни. Является ли нейрогенез автоматическим и регулярным, ведь проведение аксонов четко обнаруживается во время взрослого периода, т.к. эпителий расширяется и в нем увеличивается количество OSNs. У взрослых новые OSNs зарождаются и проецируют аксоны в луковицу. Напротив, большая часть проводимых аксонов на длинные расстояния заканчивается где-то в др. месте нервной системы взрослых. Взрослый OSN нейрогенез ставит вопрос, существуют ли механизмы, которые лежат в основе установления паттерна клубочков, поддерживают также и их сохранение. Этот вопрос попытались решить путем индукции в целом или избирательной потери из популяции OSN.
Восстановление проекций аксонов у взрослых мышей и крыс было исследовано двумя способами, которые вызывали массивные повреждения: поперечный разрез обонятельного нерва (Costanzo 2000, Christensen et al. 2001) и внутрибрюшинные инъекции dichlobenil (St John & Key 2003b). Dichlobenil разрушает избирательно OSNs не затрагивая др. типы клеток обонятельного пути, такие как обонятельные покровные клетки. При обоих методах массивная гибель OSN гарантировалась в течение нескольких дней, сопровождаемая энергичной пролиферацией клеток внутри базального слоя и дифференцировкой новых OSNs. После нескольких недель эпителий восстанавливается, но паттерн аксональных проекций не восстанавливается. У поврежденных P2 мышей (Costanzo 2000, St John & Key 2003b), меченные аксоны соединяются, но имеются многочисленные клубочки и они широко диспергированы по луковице. Сходным образом у крыс перерезка нерва нарушает распределение проекций OCAM+ и OCAM- аксонов (Christensen et al. 2001). Др. повреждающий метод заключается в пассивном вдыхании газа methyl bromide, который избирательно разрушает OSNs и поддерживающие клетки. Паттерны экспрессии всех 8 изученных ORs крыс восстанавливались спустя 3 мес. после повреждения methyl bromide (Iwema et al. 2004); проекции аксонов не изучались. Эти исследования предоставили серьезные доказательства мощной регенеративной способности обонятельного эпителия у взрослых мышей и крыс. Но они также показали, вообще-то ничего удивительного, что проведение аксонов не является нормальным после массивных повреждений.
OR-IRES-marker подход был расширен, чтобы индуцировать избирательное устранение P2
+ OSNs (Gogos et al. 2000). Генетический замысел согласуется со скрещиваниями между трансгенными мышами, несущими tet-оператор, управляющий дифтерийным токсином и gene-targeted мышами с
P2-IRES-rtTA-IRES-taulacZ аллелем. Дифтерийный токсин очень эффективен в убийстве клеток клеточно-автономным способом. Tet-оператор является минимальным промотором, который регулируется с помощью активной формы tetracycline-зависимого транскрипционного оператора rtTA. Добавление аналога tetracycline, doxycycline, к пище приводило к избирательной гибели клеток, экспрессирующих аллель
P2-IRES-rtTA-IRES-taulacZ. Предостерегает то, что в контрольных экспериментах аксоны OSNs, экспрессирующие этот P2 аллель соединяются в клубочки, которые находятся по соседству с нативными P2 клубочками. Tricistronic, двойные-IRES мутации в локусе
P2, т.о., по-видимому, изменяют существенные свойства OSNs. Индукция экспрессии дифтерийного токсина дает остаточные популяции в 2-10% от нормальных количеств, указывая тем самым, что 98-90% P2
+
OSNs убивается. Регенерация популяции P2
+ OSN ведет к появлению P2 клубочков после нескольких недель. Не удалось продемонстрировать, что аксоны от вновь возникших P2
+ OSNs формируют синапсы внутри клубочков или что эти клубочки гомогенно иннервируются аксонами от P2
+ OSNs.
EVOLUTIONARY CONSIDERATIONS
Вовлечение ORs в конвергенцию аксонов является экономичным: ORs стоят двойной цены, как распознают запахи и наводят аксоны. Эта двойная роль упрощает предназначение клубочков, если появляются новые OR гены в ходе эволюции. Если новый OR существенно отличается от любого др. OR репертуара, то клубочек для нового OR скорее всего будет новым и д. будет втиснуться куда-нибудь в паттерн клубочков. Аналогично, если два существенно отличающиеся аллеля данного OR комбинируются в индивиде благодаря скрещиванию, то д. сформироваться два сомостоятельных клубочка. Напр., полиморфизм по двум аминокислотам в M50 между линиями 129 и C57BL/6 коррелирует с самостоятельными клубочками (Feinstein & Mombaerts 2004).
Снова представление о мишенях в луковицах осложняет ограничения, накладываемые на проведение аксонов. По-видимому не разумно предполагать, что незанятые мишени в луковице уже присутствуют для новых клубочков, когда возникает новый OR ген в ходе эволюции или когда новая комбинация двух существенно отличающихся аллелей данного OR возникает при скрещиваниях. Если бы существовала позиционная информация, чтобы наводить OSN аксоны точно на мишени в луковице, то как бы эти сигналы эволюционировали совместно с громадным репертуаром OR генов? Репертуары OR млекопитающих безусловно пластичны; мыши имеют 1000-1200 OR генов, а крысы имеют 1500 OR генов.
Согласующимся заключением из всех замещений OR является то, что аксоны соединяются в клубочки в характерных позициях в луковице. Описаны замещения OR, которые предваряют образование клубочков или которые приводят к диффузным проекциям во многие клубочки. Даже экспрессия не-OR, такого как β2AR с OR локуса дает свой собственный клубочек. У трансгенных мышей с аберрантным паттерном экспрессии часто образуются новые клубочки. Эти генетические эксперименты могут воспроизводить эволюционные изменения.
ODORANT RECEPTOR CODING REGION DELETIONS
Нокаут OR гена д. стать критическим тестом роли ORs в проведении аксонов. Фенотип OR нокаутов на первый взгляд вводит в заблуждение и его интерпретация очень сильно осложнена.
Описаны 4 стандартные нокаутные мутации (как ΔOR) в P2 (Wang et al. 1998), в M71 и M72 (Feinstein et al. 2004) и в MOR28 (Shykind et al. 2004). Под нокаутом я понимаю делецию без-интронной кодирующей области, от ATG до стоп кодона. Удержание neo избирательного маркерного гена в ΔMOR28 (Shykind et al. 2004) конфликтует с превалирующими стандартами, т.к. ген neo влияет на паттерны и уровни экспрессии OR генов и многих др. генов, таких как immunoglobulin и гомеобоксные гены. Согласуется с возможным эффектом сохранения гена neo экспрессия, наблюдаемая в тканях вне обонятельной системы e ΔMOR28-IRESCre-
FNF мышей и у MOR28-IRES-Cre-FNF мышей (O'Donnell 2004). Эти внеобонятельные паттерны были охарактеризованы как неправильная экспрессия (O'Donnell 2004) и ограничили заключение о стабильности выбора OR гена, т.к. неправильная экспрессия может также возникать и внутри обонятельного эпителия. Переключение генов (Shykind et al. 2004) могло бы отражать передергивание (twitching) гена (мерцание или взрывы экспрессии OR генов), которое могло бы усиливаться при удержании гена neo в локусе MOR28 (Shykind et al. 2004).
OSNs, которые экспрессируют ΔP2 аллель, как было установлено, проецируют аксоны, которые разбредаются по поверхности луковицы и в последствии гибнут (Wang et al. 1998). Исчезновение этих OSNs было позднее интерпретировано по-новому, как угасание ΔP2 локуса (Shykind et al. 2004). Напротив, OSNs, которые экспрессируют ΔM71 или ΔM72 аллель, присутствуют в изобилии вплоть до 6 мес. (Feinstein et al. 2004). Стабильность генного выбора в M71 локусе далее была продемонстрирована отслеживанием клонов с M71-IRES-Cre мутацией, которая свободна от гена neo (Li et al. 2004). Др. объяснением различий между MOR28 и M71 находками заключается в том, что частота переключения гена или twitching гена может зависеть от OR локуса.
Несмотря на эти предостережения, фенотипы 4 ΔOR мутаций оказались информативными в отношении наведения аксонов, но способом, который не столь очевиден. OSNs, которые экспрессируют ΔOR локус, часто ко-экспрессируют др. OR локус. В одной популярной модели эта ко-экспрессия вытекает из-за отсутствия OR обусловленной негативной петли обратной связи (Serizawa et al.2003, Lewcock & Reed 2004, Shykind et al.
2004). Согласно альтернативной модели ко-экспрессия двух OR происходит обычно, хотя и не очень часто; ко-экспрессия д. быть только переносимой и совместимой с жизнеспособностью OSN, когда один OR локус (такой как ΔOR локус) не дает функционального OR (Mombaerts 2004a, Feinstein et al. 2004). Любая из моделей распознает, что ΔOR+ OSNs
ко-экспрессируют функциональные OR. Независимо от интерпретации ко-экспрессии, во всех 4-х ΔOR фенотипах и у трансгенных мышей с ΔMOR28 локусом (Serizawa et al. 2003), аксоны проецируются диффузно на луковицу, а некоторые аксоны вступают в клубочки (Feinstein et al. 2004). Эти диффузные проекции занимают широкий домен луковицы. Для ORs с дорсальной экспрессией в эпителии и дорсальными клубочками, такими как M71 и M72, этот домен оказывается дорсальным; для OR с промежуточной экспрессией в эпителии, такого как P2, этот домен также промежуточный; а для вентральных OR, таких как MOR28, этот домен вентральный.
Если бы роль OR была ограничена финальной стадией наведения аксонов, после того, как аксоны наводились с помощью механизмов, не зависящих от OR в небольшую область поверхности луковицы, то можно было бы предсказать, что аксоны от ΔOR
+ OSNs будут ограничиваться такой же небольшой областью. В качестве аргумента, не имеет отношения к делу, экспрессируется ли второй и функциональный OR в ΔOR
+ OSNs соответственно локализации OSN в эпителиальном слое; ободряет, что у ΔMOR28 трансгенных мышей ко-экспрессируемые OR гены, по-видимому, соответствуют локализации (Serizawa et al. 2003). Качественные особенности второго OR д. быть единственным известным отличием среди ΔOR
+ OSNs. Все др. молекулы наведения д. экспрессироваться в той же самой комбинации и на том же самом уровне, соответствующим мутантному локусу. (Вряд ли ORs будут дифференциально затрагивать комбинации и уровни молекул наведения). Т.о., широкие домены на луковице, которые иннервируются аксонами от ΔOR
+ OSNs отражают вклад не зависимых от OR детерминантов, a OR выполняют доминантную роль внутри этих широких доменов. Следовательно, роль OR ни в коем случае не ограничена финальным выбором мишени внутри малой области.
MOR256-17
Инициация проекций аксонов из эпителия в луковицы в раннем развитии может использовать специальный случай OR-зависимых взаимодействий.
MOR256-17 экспрессируется большой фракцией клеток обонятельного эпителия на ст. E10-E12 (Conzelmann et al. 2002; Schwarzenbacher et al. 2004, 2006). Были получены специфические антитела против MOR256-17 (Strotmann et al. 2004). Интересно, что MOR256-17 экспрессируется также клетками в ситовидной мезенхиме, которая расположена между эпителием и будущей луковицей. Эти клетки общими признаками с OSNs, но не являются типичными нейронами. Около 50% этих клеток в мезенхиме экспрессирует in the mesenchyme MOR256-17. Некоторые из клеток MOR256-17+ в мезенхиме располагаются стратегически там, где OSN соединяются вместе и образуют пучек. Аксоны от MOR256-17+ OSNs достигают луковицы к ст. E13 и окружают луковицу целиком к E18, когда они соединяются в клубочки на вентральных позициях. Эти наблюдения позволят высказать предположение, что MOR256-17+ мезенхимные клетки могут осуществлять общее наведение аксонов от эпителия к луковицам во время раннего развития; вообще-то известны некоторые др. гены ORс подобной функцией.
Между прочим, первым описанием
in situ гибридизации ORs было
MOR256-17, который был тогда назван
OR3 (Nef et al. 1992). В этом исследовании сообщалось о временном появлении OR3
+ клеток в ситовидной мезенхиме мышей. На постнатальный день 10, ~1000 OR3
+ OSNs было вычислено в эпителии. Др. группа подсчитала сходное число на ст. E14 (Schwarzenbacher et al. 2004). Т.о., эта популяция OSNs, по-видимому, резко возрастает между E10 и E14 и затем стабилизируется, Ранняя экспансия популяции MOR256-17
+и экспрессия этого гена в мезенхимных клетках являются двумя показателями специальной функции в инициации проекций аксонов. Если эта специальная функция зависит от самого MOR256-17, то вообще-то этот OR д. быть необычно хорош при взаимодействии со многими др. ORs, или вообще он д. быть столь хорош на этих ранних стадиях из-за рыхлого ограничения взаимодействий.
THE OCAM SAGA
В общем топографическая связность может быть выявлена в проекциях аксонов от эпителия к луковицам в основном вдоль дорсо-вентральной оси. В последние 20 лет паттерн пространственной и временной экспрессии Olfactory Cell Adhesion Molecule (OCAM) стал ведущим кандидатом на роль молекулярного детерминанта для общей топографической коммуникабельности. Но надежда увяла, когда был проанализирован нокаутный OCAM фенотип (Walz et al. 2006a). Сага об OCAM д. служить предупреждением приписывания существенной роли молекулам в проведении аксонов на базе интересных паттернов пространственной и временной экспрессии.
OCAM является членом сверхсемейства immunoglobulin (Alenius & Bohm 1997, Yoshihara et al. 1997) и тесно связан по последовательностям с Neural Cell Adhesion Molecule (NCAM) и fasciclin II. OCAM осуществляет гомофильные взаимодействия in vitro, но не соединяется с NCAM (Yoshihara et al. 1997). Экспрессия OCAM в эпителии ограничена OSNs в вентральных регионах, которые проецируют свои аксоны в клубочки вентро-латаральных частей луковиц. OCAM временно экспрессируется в дендритах mitral/tufted клеток в дорсо-медиальном аспекте луковиц крыс, формируя зеркальное отражение проекций OCAM- аксонов (Treloar
et al. 2003). OCAM сохраняется на OSNs у взрослых, но исчезает из mitral/tufted клеток вскоре после рождения.
Неутешительно, что при различных измерениях OCAM нокаутов не было выявлено очевидного эффекта на топографию проекций в основной и добавочной обонятельной системах (Walz et al. 2006a). Скорее нарушена организация внутри клубочков: Сегрегация аксон-дендрит и дендрит-дендрит синаптических circuits внутри клубочков оказалась менее выраженной. Т.о., с момента открытия, когда было предположено, что OCAM выполняет функции, связанные с проведением аксонов, в частности с общей топографической коммуникабельностью, фенотип нокаутов заставил )временно?) отказаться от этой гипотезы. Аргументы о перекрывании без сомнения были предложены: др. молекула(ы) д. компенсировать потерю OCAM. Но эти альтернативные гипотезы выдвигались многие годы, но стало ясно, что стоит изучать неперекрывающиеся механизмы. Сегодня OCAM является удобным маркером для (вентральных) OSNs, которые не желают смешиваться с (дорсальными) OCAM
- OSNs (Chehrehasa et al. 2005), но OCAM сам по себе не отвечает на это дифференциальное смешивание.
EPHRINS AND EPHRIN RECEPTORS
Роль ephrins и их рецепторов в retinotectal или в retinocollicular проекциях описана в деталях (Lemke & Reber 2005). В зрительной системе слой периферических рецепторов проецируется в высшие области головного мозга, так что сохраняются сходные взаимоотношения. Позиционные координаты Cartesian x-y типа картированы от одного двухмерного слоя нейронов к соседнему двухмерному слою нейронов с которыми они образуют синапсы. Распределение ephrin лигандов и Eph рецепторных тирозин киназ организовано градированным и комплементарным образом вдоль ортогональных осей сетчатки и тектума. Напротив проекции аксонов от обонятельного эпителия к луковицам следуют совершенно противоположным принципам организации: OSNs, которые разбросаны по большой области эпителия проецируют аксоны в небольшие клубочки в луковице. Из первого принципа неясно если да, то каковы механизмы, поддерживаемые с помощью ephrins и Eph рецепторов в зрительной системе могут быть распространены на обонятельную систему.
Ephrins и Eph рецепторы экспрессируются в виде высоко сходных пространственного и временного паттернов, как OSNs, так и клетками луковиц (St John et al. 2002b). Из-за множества взаимодействий между ephrins и Eph
рецепторами, не возможно прямо предсказать функциональный исход их взаимодействий - и нокаутов. Отсутствуют очевидные градиенты экспрессии в эпителии и луковице. Уровни Ephin-A обнаруживают мозаичные паттерны в гломерулярном слое, что лишено очевидной логики: клубочки, экспрессирующие высокие уровни, соседствуют с клубочками, экспрессирующими низкие уровни (Cutfortht et al. 2003).
Функциональные доказательства роли ephrins и Eph рецепторов в проведении аксонов в обонятельной системе ограничены одним сообщением (Cutforth et al. 2003). У мышей с нокаутами ephrin-A3 и ephrin-A5, позиции клубочков P2 и SR1 подвергаются сдвигу назад. SR1 является типичным OR и экспрессируется в 50% OSNs septal органа (Tian & Ma 2004), малого поселения, отдаленного от основного обонятельного эпителия; он не является репрезентативным для репертуара OR. Абсолютная позиция клубочков трудна для определения из-за возможных различий в размере и форме луковиц у мутантных мышей. Интересно, что др. ephrin, ephrin-A2, оказывает негативный эффект на пролиферацию нервных предшественников и на нейрогенез, так что нокаутные по ephrin-A2 мыши имеют больше интернейронов в луковицах (Holmberg et al. 2005). Сходным образом, незначительные изменения в размерах или форме луковиц у двойных мутантов
ephrin-A3/ephrin-A5 могут вызывать кажущееся смещение позиции клубочков, которые могут не отражать непосредственно роли этих ephrins в совместном предопределении их позиций. Обратный эксперимент с избыточной экспрессией ephrin-A5 в P2
+ нейронах посредством
P2-IRES-ephrin-A5-IREStaulacZ мутации (Cutforth et al. 2003), страдает от того же самого возражения, т.к. эксперименты с участием
P2-IRES-rtTA-IRES-taulacZ мутации (Gogos et al. 2000), которые дают новые клубочки, которые отличны от нормальных P2 клубочков. Tricistronic мутации в локусе
P2 могут влиять на положение клубочков независимо от того, какие др. белки ко-экспрессируются, вообще-то за счет уменьшения количества продуцируемого P2 белка. Из дальнейших касательств, anteriorization P2 клубочков с помощью избыточной экспрессии ephrin-A5 происходит в пределах только одного из двух клубочков и наблюдается только в 60% случаев. Из-за очевидных ограничений этого исследования (Cutforth et al. 2003), роль ephrins и Eph receptors в управлении образованием топологической карты в луковице остается спорной.
SEMAPHORIN 3A
Несколько линий нокаутных мышей было изучено в отношении аномалии проведения аксонов в обонятельной системе. В большинстве случаев довольно приближенные показатели, такие как окрашивание лектинами DBA, .skb использованы для определения исхода (Treloar et al. 1997). Более сфокусированный подход - это изучение специфических клубочков у OR-IRES-marker мышей. P2 клубочки привлекли большое внимание, но большинство результатов негативны. P2 клубочки оказались нечувствительны к отсутствию CNGA2 (Lin et al. 2000, Zheng et al. 2000), Tbr-1 и Dlx-1/Dlx-2 (Bulfone et al. 1998), Golf (Belluscio et al. 1998), neutrophins и нейротрофиновых рецепторов (Nef et al. 2001) и OCAM (Walz et al. 2006a). Предварительные данные показали, что P2 клубочки не сохраняются у старых Golf нокаутных мышей (Belluscio 1998).
Расхождения возникли, когда две группы исследовали P2 клубочки у
Semaphorin 3A (Sema3A) нокаутных мышей. Sema3A является ассоциированным с мембранами секретируемым белком со свойствами хеморепелента для neuropilin-1-positive (npn-1
+) аксонов. Sema3A экспрессируется несколькими типами клеток в обонятельной системе, особенно обонятельными оболочечными клетками в нервном слое вентральных частей луковиц. У
Sema3A нокаутных мышей, npn-1
+ OSN аксоны неправильно идут в вентральные клубочки у новорожденных (Schwarting
et al. 2000) и у взрослых (Taniguchi et al. 2003, Schwarting et al. 2004). Schwarting et al. (2004) описывают множественные, более мелкие, неправильной формы и вентрализованные P2 клубочки у Sema3A нокаутных мышей; Taniguchi et al. (2003) не выявили различий. Нет очевидных объяснений, таких как линейные или генетического фона различия, которые бы объясняли эти расхождения. Однако, генетический фон в gene-targeted линиях мышей может подвергаться огромным изменениям между лабораториями, что мешает воспроизводимости результатов. Обычный генетический фон gene-targeted мышей 129 х C57BL/6 смешанный. Обычно, очень немногие мыши перемещаются из одной лаб. в др. (генетическое узкое горло) и колонии часто поддерживаются за счет скрещиваний между братьями сестрами (инбридинг и генетический дрейф).
Результаты могут в дальнейшем затрагиваться различиями в запах окружающей среды, в которой пребывают животные. Такие незначительные эффекты становятся важными, когда фенотип нокаутов также едва обнаружим. Решением является увеличение генетической чистоты за счет возвратных скрещиваний OR-меченных мышей и повторов в параллельных экспериментах, производимых в др. лабораториях и ли в той же лаб. позднее.
GLYCOCODE
Различные углеводы клеточной поверхности экспрессируются на OSN аксонах в виде характерных паттернов, которые могут быть выявлены путем окрашивания растительными лектинами, такими как DBA или UEA. В наружном нервном слое DBA+ аксоны отсортировываются от др. аксонов и собираются в дискретные пучки (Key & Akeson 1993). Такие углеводы клеточной поверхности, как полагают, взаимодействуют с белками, связывающими углеводы, такими как galectin-1, который экспрессируется многими клетками в обонятельной системе. Нокауты по galectin-1 дают аномалии в паттерне клубочков: DBA+ клубочки исчезают из дорсо-каудальной области луковиц (Puche et al.1996).
Гипотеза гликокода предполагает, что многочисленные углеводы клеточной поверхности предоставляют клеточной поверхности сигнатуру, благодаря которой аксоны сортируются перед OR-зависимым соединением в клубочки (St John & Key 2005). Трудно представить, как гликокод скоординированно экспрессируется среди OSNs, экспрессирующих один и тот же OR или группы ORs. Как и в случае ephrin-A (Cutforth et al. 2003), распределение DBA
+ клубочков в луковице мозаично и лишено очевидной организации: клубочки, в которых все аксоны являются DBA
+, соседствуют с клубочками, в которых лишь немногие аксоны DBA
+ и с клубочками, не содержащими аксонов DBA
+ (Lipscomb et al. 2003). Основным препятствием в прогресс в этом направлении является отсутствие точных инструментов, с помощью которых может изучаться роль углеводов клеточной поверхности. Кроме того, эти молекулы часто экспрессируются слишком многими типами клеток, чтобы можно было интерпретировать результаты. Нокауты по гену, кодирующему β-1,3-N-acetylglucosaminyltransferase 1, ключевой энзим в синтезе lactosamine glycan, предотвращают образование P2, I7 и M72 клубочков
glomeruli (Henion et al. 2005).
ACTIVITY-DEPENDENT MECHANISMS
В др. сенсорных системах, таких как зрительная система, формирование проекций аксонов во время развития подразделено на ранней фазе на зависимые от активности процессы, сопровождаемые утонченными зависимыми от активности процессами. Активности посредством нейрональной активности, обычно электрической активности, которая может быть спонтанной или разбуженной стимулами, скоррелированы или не скоррелированы. Организация проекций аксонов OSN ведет к значительному a priori скептицизму относительно роли зависимых от активности механизмов. Литература по этому вопросу бедна и противоречива. Блокирование передачи обонятельных сигналов путем нокаута субъединицы cyclic nucleotide-gated channel, CNGA2 (первоначально названного OCNC1), затрагивает проведение аксонов в некоторых ситуациях. В то время как P2 клубочки формируются нормально у CNGA2 нокаутных мышей, клубочки M72 нет (Lin et al. 2000, Zheng et al. 2000). Мозаичное сосуществование M72+ OSNs , которые экспрессируют или нет CNGA2, было осуществлено посредством извлечения выгоды из моноаллельной экспрессии OR генов; проекции аксонов от
CNGA2+ and CNGA2- OSNs рассортировываются в разные клубочки (Zheng et al. 2000). Сходная мозаичная ситуация была создана, но глобально в OSNs, базируясь на мозаичной X-инактивации у самок гена CNGA2, расположенного Х хромосоме (Zhao&Reed 2001). У молодых самок CNGA2+ и CNGA2- OSNs сосуществуют в одинаковых количествах, но со временем клетки CNGA2- исчезают, а небольшие количества клубочков, инактивированные за счет CNGA2- OSNs, сохраняются у старых мышей. Постоянное и одностороннее хирургическое закрытие одной из ноздрей увеличивает количество клубочков, иннервируемых с помощью CNGA2- OSNs. Множественные и гетерогенные M71 и M72 клубочки появляются временно во время постнатального развития (Zou et al. 2004). Эти гетерогенные клубочки исчезают обычно спустя 8 недель, но сохраняются после закрытия ноздри. Эта хирургическая процедура снижает все активности, ассоциированные с прохождением воздуха чрез носовую полость: биохимические, электрические и токсические. Увеличение жизнеспособности OSNs с закрытой стороны указывает на то, что избирательная гибель OSNs является клеточным механизмом для ремоделирования клубочков. Обонятельное обучение в вызывающем отвращение conditioning протоколе ускоряет ремоделирование клубочков (Kerr & Belluscio 2006). Однако, закрытие ноздри и скрещивание с CGNA2 нокутными мышами не вмешивается во внутриклубочковую сегрегацию проекций аксонов, наблюдаемую в M71-M72 гибридных скрещиваниях (Feinstein & Mombaerts 2004): эти характерные фенотипы, по-видимому, являются первичным эффектом ORs. Условная экспрессия легкой цепи tetanus toxin, которая ингибирует высвобождение синаптических нейротрансмиттеров, вмешивается в поддержание P2 клубочков, когда избирательно экспрессируется в P2+ OSNs, но не когда широко экспрессируется в зрелых OSNs (Yu et al. 2004). Условная избыточная экспрессия внутрь очищающих калиевых каналов Kir2.1 приводит к драматическому снижению спонтанного возбуждения и предотвращает образование P2 и MOR23 клубочков, в то время как MOR28 клубочки оказываются неожиданно нормальными. Т.к. экспрессия tetanus легкой цепи и Kir2.1 блокируют как спонтанную, так и вызванную запахом активность, то дифференциальный анализ вкладов этих двух типов активностей не может быть проведен непосредственно. Изучение главной роли вызванной запахом и скоррелированными паттернами активности ещё более осложнено гетерогенностью реакций на запахи OSNs, которые экспрессируют один и тот же OR (Bozza et al. 2002, Grosmaitre et al. 2006). Напр., порог реакции на пахучий лиганд lyral варьирует в 100 раз в MOR23+ OSNs. Если эти наблюдения можно экстраполировать на ситуацию in vivo, то паттерны активности могут варьировать существенно среди OSNs, экспрессирующих один и тот же OR.
На ум приходит единственное, что активность д. играть некоторую роль в проведении аксонов в обонятельной системе, но такая роль не может быть инструктивной и вообще не столь интересна. После этого, почему активность не может играть роль в модуляции фенотипов клеток, которые специализированы в обеспечении паттернов активности? OSN, которые не могут отвечать на запахи или не могут продуцировать потенциалы действия д.быть нефункциональными по многим причинам.
EFFICIENCY OF AXONAL WIRING
Всепроникающая и слепая вера в литературе, что OSNs являются безупречными клетками. Они д. управлять так, чтобы экспрессировать один, и только один, функциональный OR ген с эффективностью в 100%, чему помогает обратная петля связи и их аксоны д. находить свои клубочки с эффективностью 100%. Итог и в самом деле кажется безупречным: 100% аксонов, иннервирующих M72 клубочки, сформированы M72+ OSNs (Treloar et al. 2002). Но процесс не нужно путать с итогом; д. быть существенное изнашивание OSNs на множественных стадиях.
Картины конвергенции аксонов OR меченных мышей могут быть обманчивы. Они не показывают всех аксонов, которые исходят от OSNs, экспрессирующих этот OR, заканчивающихся стабильно в этих клубочках. Вообще-то только те аксоны, которые иннервируют правильно клубочки, сохраняются, а OSN с неправильным проведением аксонов не выживают. Картины конвергенции аксонов могут скрывать существенный уровень неэффективно проведенных аксонов. То что выглядит как величественный подвиг проведения аксонов может быть продуктом безжалостного отбора, опустошающего большинство OSNs так что сохраняется только незначительное меньшинство. Контекстуальная модель само-сортировки также скорее всего не защищена от ошибок и даже не столь уж эффективна: конвергенция аксонов д. происходить в основном в результате случайности, а не в результате прямого наведения на цель.
CONCLUSION
There are other views on axonal wiring. For many authors, it remains irresistible to hypothesize spatial cues in the bulb that would specify the positions for each and every glomerulus. Key & St John (2002),
St John et al. (2002), Axel (2005), Nedelec et al. (2005), and Komiyama & Luo (2006) present interesting reviews and opinions. The
growth of knowledge depends entirely on the existence of disagreement (Popper 1994).
I admit to a bias in my review, in which I
interpret the literature through the perspective of our contextual model of self-sorting and by dismissing targets in the bulb. But this
model reduces the problem to the essence: OR-specific axonal convergence is OR dependent. The model is parsimonious and does not
invoke complex scenarios with multiple layers of guidance control that would have to act in concert with the expressed OR. The model
is attractive from an evolutionary viewpoint: The bulb is receptive to experiments of nature,and coevolution of genes for guidance
factors and OR genes is not required. The challenge for the model is unambiguous:How can a multitude of axonal interactions comparing
each other for OR similarity generate an array of 1600–1800 glomeruli at positions that are so reproducible?
I do not disregard the potential contributions
of ephrins and ephrin receptors, adhesion molecules, cell surface carbohydrates, activity-dependent mechanisms, and so forth.
But there are other, unrelated wiring problems in the bulb, for example, projections of mitral/tufted cell dendrites to glomeruli, connectivity
of interneurons, and centrifugal projections. Molecules with intriguing expression patterns (Lin et al. 2004) or knockout phenotypes
may be involved in these other wiring problems and only indirectly and secondarily in the wiring of OSN axons.
Research on axonal wiring in the olfactory system ought to be focused primarily on explaining how ORs function in axonal wiring.
This problem is well defined but remains mysterious. Its solution may require a patient continuation of the genetic approach combined
with experiments in biophysics.
SUMMARY POINTS
|
|
1. The mouse genome has?1000 intactORgenes. EachOSNexpresses just one allele of oneORgene. Axons of OSNs that express the sameORproject to the same glomeruli: typically to one glomerulus per half-bulb. For each OR, there are thus at least four glomeruli per mouse.
2. The expressed OR determines the glomerulus that is innervated: Altering the OR oding region by targeted mutagenesis alters the position of the glomerulus that is formed. The first evidence for the role of the OR in axonal wiring was provided in 1996, but no mechanistic insights as to how ORs function precisely in axonal wiring have emerged.
3. OSNs axons do not converge onto a glomerulus but coalesce into a glomerulus. Axons do not project to a preexisting structure, but they dynamically form a glomerulus by coming together and forming synapses with other types of neurons.
4. The glomerular array is not a stereotyped spatial map because the OR identity of a particular glomerulus cannot be derived from its position, shape, or size. There is no evidence for targets in the olfactory bulb to which axons would be guided.
5. An in vivo site-directed mutagenesis study has led to a contextual model of self-sorting. OSNaxons would sort themselves out into glomeruli through sampling of otherOSN axons with their growth cones. This self-sorting would be mediated by hypothetical
homophilic interactions between complexes that contain ORs or OR fragments.
6. The ~1800 glomeruli are not wired simultaneously but emerge over a period of a few days perinatally. This developmental asynchrony may be instrumental in axonal wiring. Insufficient attention has been paid to temporal control.
7. Once established, the glomerular array must be maintained in the face of OSN death to preserve the constancy of olfactory perception. Like other cells in epithelia, OSNs are turning over throughout the lifetime of the organism. OSN renewal poses an additional challenge to the mechanisms of axonal wiring.
8. A pervasive and implicit belief in the literature is that the processes of OR gene choice and axonal wiring are perfect. Quite the opposite may be the case: There may be substantial attrition of OSNs at various stages. What appears as a magnificent feat
of axon guidance may be the product of merciless selection that wastes many OSNs and perhaps only retains a small minority.
|
Сайт создан в системе
uCoz