Three developmental compartments involved in rib formation  YOKO MIZUTANI-KOSEKI and HARUHIKO KOSEKI YOKO MIZUTANI-KOSEKI and HARUHIKO KOSEKIInt. J. Dev. Biol. 49: 325-333 (2005) doi: 10.1387/ijdb.041932ha | |
|
When the thoracic somitic mesoderm was separated from the neural tube and the notochord with a piece of aluminum foil in two-day chick embryos, seven days after the operation ribs lacked their proximal part. The embryos were rescued by co-transplanting the notochord, the
ventral half of neural tube, or QT6 cells transformed with Shh, on the somite side of the aluminum foil insert. Thus, proximal rib development depends on the notochord and the ventral neural tube,
an effect which might be mediated through Shh secreted by these axial tissues. On the other hand, when the thoracic somitic mesoderm was separated from the surface ectoderm by a piece of
polyethylene terephthalate film, the distal parts of the ribs were missing, suggesting that distal rib development depends on surface ectoderm. In these embryos, expression of Pax3 was weak and
perturbed showing that the dermomyotome developed abnormally. It is not clear whether the development of distal rib is mediated by the dermomyotome, or the ectoderm. It has previously
been shown that sternal rib development depends on lateral plate mesoderm. As to the distal rib, it is considered to be composed of two parts. Thus, the rib is composed of three developmental compartments, in agreement with a recently presented classification of somite derivatives as
primaxial and abaxial.
|
From the neural crest to the somite Т.к. сегментация нервного гребня обусловливается взаимодействием с сомитами (Rickmann et al., 1985; Goldstein et al., 1990), то сегментация сомитов может играть фундаментальную роль. Далее клетки обеих тканей мигрируют от своего первоначального места детерминации к финальной позиции, когда они дифференцируются, чтобы сформировать структуры, к которым они предназначены. Наконец, что очень важно, их онтогенетический репертуар варьирует вдоль ростро-каудальной оси. Краниальная часть нервного гребня может дифференцироваться в хрящ, тогда как туловищная часть гребня нет. Сомиты формируют регион-специфические единицы осевого скелета. Напр., в осевом скелете птиц и млекопитающих рёбра обнаруживаются исключительно в торакальной области. Rib formation Недавние находки в биологии развития пролили новый свет на механизмы эволюции (Carroll et al., 2001). Что касается филогении позвоночных, то осевой скелет служил одним из главных предметом исследования (Burke et al., 1995). Ребро является простой и важной моделью для изучения, как региональной спецификации развивающихся позвоночных, так и разнообразия животных. Разнообразие ребер вдоль оси тела позвоночных является не только онтогенетическим признаком, но также филогенетическим и находится под контролем Hox генов (Burke et al., 1995). Хотя ребра и ограничены торакальной областью у птиц и млекопитающих, однако, известен их вклад в некоторые позвонки, например костальные (costal) отростки поясничных позвонков (Standring, 2005).
Ребра, также как и позвонки, как известно, происходят из сомитов (Seno, 1961; Pinot, 1969; Christ et al., 1974; Chevallier, 1975; reviewed by Christ and Wilting, 1992; Tam and Trainor, 1994; Christ et al., 1998). Регион-специфический морфогенез осевого скелета может быть подразделен на две фазы. В первой фазе сомитная мезодерма регионализуется. Kieny et al. (1972) впервые показали с помощью гетротопических трансплантаций, что сомиты и сегментная (segmental) пластинка у эмбрионов кур уже регионализованы. Будут ли они формировать ребра или нет зависит от их исходной позиции вдоль ростро-каудальной оси эмбриона, эти находки подтверждены Chevallier (1975) с помощью курица/перепел химер. Если торакальные сомиты или сегментная пластинка трансплантируются в шейную область, то они образуют рёбра и, если шейная сомитная мезодерма трансплантируется в торакальную область, то она не даёт там ребер. Наши данные подтверждают, что такая регионализация происходит в презумптивной сомитной мезодерме, начиная со стадии первичной полоски (Sakamoto and
Aoyama, in preparation).
Вторая фаза является последовательным процессом дифференцировки и морфогенеза сомитных клеток, которые конструируют морфологию скелета. Когда сомит оказывается отшнурованным от сегментной пластинки, то он состоит из эпителиального мешка, заключающего мезенхимные клетки сомитоцеля. После этого сомит дифференцируется в две части, вентро-медиальный мезенхимный склеротом и дорсо-латеральный эпителиальный дермомиотом. Эти изменения в эпителиальном сомите зависят от окружающей ткани (Gallera, 1966; Swalla and Solursh, 1984; Aoyama & Asamoto, 1988; Christ et al., 1992; Aoyama, 1993; Dietrich et al., 1997; также rev Dockter, 2000; Christ et al., 2004) и инициируются в течение 3-х ч (Aoyama & Asamoto, 1988), хотя склеротомные клетки, будучи дифференцированными, всё ещё могут изменять свою онтогенетическую судьбу (Dockter and
Ordahl, 2000). Дифференцировка склеротома зависит от аксиальных тканей, таких как хорда и донная пластинка, и от Shh, продуцируемого этими тканями (Watterson et al., 1954; Teillet and Le Douarin, 1983; Brand-Saberi et al., 1993; Fan and Tessier-Lavigne, 1994; Johnson et al., 1994; Fan et al., 1995; Chiang et al., 1996; Borycki et al., 1997; Dietrich et al., 1997). С др. стороны, дифференцировка дермомиотома зависит от поверхностной эктодермы (Gallera, 1966; Dietrich et al., 1997). Далее, миотом появляется на медиальном и латеральном краях дермомиотома. Первый даёт epaxial мышцы, а последний hypaxial мышцы. Развитие epaxial мышц рассматривается как зависимое от хорды и нервной трубки, а hypaxial мышц от мезодермы латеральной пластинки и поверхностной эктодермы (rev. Christ and Ordahl, 1995; Dietrich, 1999; Borycki and Emerson, 2000). Т.о., ткани, окружающие сомит регулируют спецификацию сомита на компартменты.
В данном исследовании мы изучали роль тканей, окружающих сомит, в формировании ребер. Когда взаимодействие между сомитом и аксиальной тканью прерывается путем вставления кусочка непроницаемого фильтра между тканями у 2-дневных эмбрионов кур, то проксимальные части ребер не развиваются, тогда как прекращение взаимодействия между поверхностной эктодермой и сомитом вызывает потерю дистальной части ребер. Ранее мы показали, что латеральная пластинка существенна для развития грудинной (sternal) части ребер у эмбрионов птиц (Sudo et al., 2001). Здесь мы показали, что три части ребер проксимальная, дистальная и стернальная являются онтогенетически отличающимися компартментами. Мы рассматриваем эти компартменты с связи с новой классификацией производных сомитов, primaxial и abaxial (Burke and Nowicki, 2003).
Results Rrole of medial tissues in the development of the proximal rib
Block of somite-axial tissue interaction
A piece of aluminum foil of about 1/4-1/3 mm by 1 mm was inserted
between the thoracic somitic mesoderm along 6 somites and the
neural tube and notochord in a 2-day chick embryo (Fig.1A, B). The
somitic mesoderm concerned was at different levels, i.e., at different
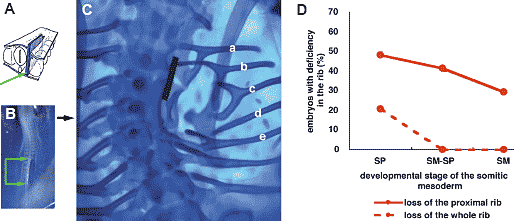 Fig. 1. Deficiency in the rib caused by blocking the interaction between the thoracic somitic mesoderm and the axial tissues in 2-day chick embryos. (A) Schematic drawing of the operation. A piece of aluminum foil (ca. 1/4-1/3 mm X 1 mm; blue rectangle, indicated with green arrow) was inserted between the axial tissues (the neural tube and the notochord) and the somitic mesoderm. (B) Dorsal view of the embryo immediatly after the operation. Position of the aluminum foil insert is indicated with green arrows. (C) Whole mount Alcian blue staining showing the cartilaginous skeleton 7 days after the operation. Ribs display various degrees of malformation in their proximal part (a-e; detail in text). In the most severe cases, the proximal rib (head, neck and tuberculum of rib) is completely lost (d). (D) Incidence of embryos with rib deficiency depends on the developmental stage of the somitic mesoderm at the operation, i.e., unsegmented segmental plate (SP), somitic mesoderm including both segmented somites and unsegmented segmental plate (SM-SP) or already segmented somites (SM). The earlier the developmental stage of somitic mesoderm operated, the more frequently the proximal rib is missing (red solid line). Only when the barrier was inserted between the segmental plate and the axial tissues, was the whole rib missing (red broken line). Fig. 1. Deficiency in the rib caused by blocking the interaction between the thoracic somitic mesoderm and the axial tissues in 2-day chick embryos. (A) Schematic drawing of the operation. A piece of aluminum foil (ca. 1/4-1/3 mm X 1 mm; blue rectangle, indicated with green arrow) was inserted between the axial tissues (the neural tube and the notochord) and the somitic mesoderm. (B) Dorsal view of the embryo immediatly after the operation. Position of the aluminum foil insert is indicated with green arrows. (C) Whole mount Alcian blue staining showing the cartilaginous skeleton 7 days after the operation. Ribs display various degrees of malformation in their proximal part (a-e; detail in text). In the most severe cases, the proximal rib (head, neck and tuberculum of rib) is completely lost (d). (D) Incidence of embryos with rib deficiency depends on the developmental stage of the somitic mesoderm at the operation, i.e., unsegmented segmental plate (SP), somitic mesoderm including both segmented somites and unsegmented segmental plate (SM-SP) or already segmented somites (SM). The earlier the developmental stage of somitic mesoderm operated, the more frequently the proximal rib is missing (red solid line). Only when the barrier was inserted between the segmental plate and the axial tissues, was the whole rib missing (red broken line). stages of somite development. The foil piece was inserted at the level of either unsegmented thoracic somitic mesoderm (segmental plate, SP), either both somites and segmental plate (SM-SP), or already segmented somites (SM). Seven days after the operation, the proximal ribs displayed various malformations. Fig.1C shows a typical embryo with various malformations of proximal ribs. Compared to a more or less normal rib (a), the tuberculum (b), the head (e) or both were separated from the vertebral column and in more severe cases the proximal rib was completely missing (d). Such malformations are not caused by the toxicity of aluminum, because, in seven cases, insertion of smaller pieces of aluminum (0.2-0.3 mm square) did not affect formation (data not shown). The appearance of a deficiency in the proximal rib was found to
depend on the developmental stage of the somitic mesoderm when
separated from the neural tube and notochord (Fig.1D). 14 out 29
embryos (48%) in the case of non-segmented thoracic somitic
mesoderm (stage SP in Fig.1D abscissa), 12/29 (41%) in the case of
both segmental plate and somites (stage SM-SP in Fig.1D abscissa),
17/5 (29%) in the case of already segmented somites (stage SM in
Fig.1D abscissa), respectively (Fig.1D, solid line).
When the piece of foil was inserted between the thoracic segmental
plate and the axial tissues, a whole rib was often lost (6 out 29
embryos = 21%) (Fig.1D, dotted line). The whole rib was never lost
in the case of operations at the SM-SP (n=29) or SM stages (n=17). Pax1 expression pattern Koseki et al. (1993) showed that Pax1 mediates of the notochordal
signal which controls the ventral vertebra and the proximal rib. To
know whether the block of the axial tissue affects Pax1 expression as
well as proximal rib development, the expression pattern in operated
embryos was examined by whole mount in situ hybridization using
a digoxigenin labeled riboprobe for quail Pax1 cDNA.
Two days after the operation, the expression of Pax1 in the
somites was down regulated at the site where the aluminum foil was
inserted (Fig.2A, B; between two green arrows) compared to those
on the opposite non-operated side. Elsewhere, Pax1 was expressed
equally on the operated and on the non-operated side. Back-transplantation of axial tissues
Finding that blocking the interaction between the somite and the
axial tissues caused a malformation in the proximal rib suggests that
proximal rib development depends on the neural tube and/or the
notochord. However, it is not clear which tissue is required to form the
proximal rib. To clarify this issue, after inserting a piece of aluminum
foil between the axial tissues and the somitic mesoderm, we inserted
either the dorsal half of neural tube, the ventral half of neural tube, or
the notochord from quail embryos between the foil and the somitic
mesoderm (Fig.3A).
Seven days after back-transplantation of the notochord, the
ventral neural tube or the dorsal neural tube at stage SP, the ratio of
embryos missing proximal ribs to the total number of the operated
embryos were 4/13 (31%), 4/10 (40%) and 9/16 (56%), respectively
(Fig.3B). They were 2/18 (11%), 1/15 (7%) and 13/20 (65%),
respectively, in cases operated at stage SM-SP and 0/6 (0%), 0/9
(0%) and 2/5 (40%), respectively, in cases operated at stage SM.
When the aluminum foil was inserted without any tissues, the ratio of
embryos missing proximal ribs were 12/29 (41%), 5/17 (29%) and 14/
29 (48%) as stated above. Thus the incidence of the deficiency in the
proximal rib was lower in embryos with back transplanted notochord
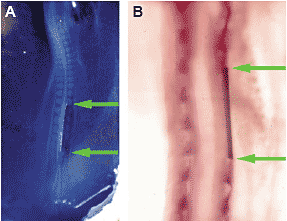 Fig. 2. Pax1 expression pattern in embryos when interaction between the somitic mesoderm and the axial tissues is blocked. (A) Dorsal view of 2-day chick embryo immediatly after the operation. Position of the
aluminum insert is indicated with two green arrows. (B) Whole mount in situ
hybridization using Pax1 probe performed of the embryo 2 days after the
operation. Dorsal view. Upper is rostral. Pax1 expression is not found in the somites that are separated from the axial tissues with the aluminum barrier (between two green arrows), while a periodical Pax1 expression pattern was seen on the opposite, non-operated side. Rostral to and caudal to the operated site, Pax1 was expressed on both sides concordantly. Fig. 2. Pax1 expression pattern in embryos when interaction between the somitic mesoderm and the axial tissues is blocked. (A) Dorsal view of 2-day chick embryo immediatly after the operation. Position of the
aluminum insert is indicated with two green arrows. (B) Whole mount in situ
hybridization using Pax1 probe performed of the embryo 2 days after the
operation. Dorsal view. Upper is rostral. Pax1 expression is not found in the somites that are separated from the axial tissues with the aluminum barrier (between two green arrows), while a periodical Pax1 expression pattern was seen on the opposite, non-operated side. Rostral to and caudal to the operated site, Pax1 was expressed on both sides concordantly. or ventral neural tube and was slightly higher in the embryos with dorsal neural tube transplants than in the embryos without backtransplantation. Thus, back-transplantation of the notochord or the ventral neural
tube rescued the embryo from the deficiency in the proximal ribs,
while the dorsal neural tube did not. Rather, the dorsal neural tube
appeared to promote the deficiency. These findings suggest that the
notochord and/or the ventral neural tube are responsible for proximal
rib formation. Co-transplantation of Shh-producing cells It is well known that the notochord and the floor plate, that is, the
ventral most part of the neural tube, induce motor neurons (Yamada
et al., 1991; Roelink et al., 1995) and sclerotomal differentiation (Fan
and Tessier-Lavigne, 1994; Johnson et al., 1994; Fan et al., 1995) by
producing and secreting Shh. To know whether the dependence of
the proximal rib development on the notochord and/or the ventral
neural tube is also mediated by Shh, we examined the effect of
transplanting Shh-producing cells. After inserting a piece of aluminum
foil between the axial tissues and the thoracic somitic mesoderm at
SM-SP stage, QT6 cells transformed with Shh gene were transplanted
between the foil and the somitic mesoderm (Fig.4A). As control, we
used the original QT6 cells instead of their transformants (Fig.4B).
Seven days after the operation with the transplantation of Shhproducing
QT6 cells or of the original QT6 cells, the ratio of embryos
missing proximal ribs were 3/18 (17%) and 8/12 (67%), respectively.
The incidence of the proximal rib deficiency was thus much lower in
the embryos inserted with barrier and Shh-producing cells than in
those inserted with barrier and Shh-non-producing cells.
Thus the Shh producing QT6 cells rescued embryos in which the
interaction between the axial tissues and the thoracic somitic
mesoderm was blocked, so that proximal ribs were normal (Fig.4C),
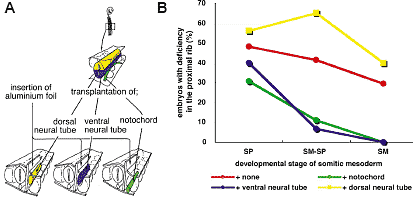 Fig. 3. Back-transplantation experiments
showing the part of axial tissues responsible
for proximal rib morphogenesis. (A) Schematic
drawing of the experiment. Insertion of a piece of
aluminum foil between the somitic mesoderm and
the axial tissues of 2 day chick embryos was
followed by transplantation of the dorsal half of
neural tube, the ventral half of neural tube, or the
notochord from quail embryos, between the
aluminum insert and the somitic mesoderm. (B)
Incidence of embryos with a deficiency in the
proximal rib 7 days after the operation. Compared
to the control embryo with only an aluminum insert (red line), the notochord (green line) and ventral half of neural tube (blue line) have rescued the embryos from the deficiency at least when they have been
operated at SM-SP or SM stage, while the dorsal half of neural tube promoted the malformation (yellow line). Fig. 3. Back-transplantation experiments
showing the part of axial tissues responsible
for proximal rib morphogenesis. (A) Schematic
drawing of the experiment. Insertion of a piece of
aluminum foil between the somitic mesoderm and
the axial tissues of 2 day chick embryos was
followed by transplantation of the dorsal half of
neural tube, the ventral half of neural tube, or the
notochord from quail embryos, between the
aluminum insert and the somitic mesoderm. (B)
Incidence of embryos with a deficiency in the
proximal rib 7 days after the operation. Compared
to the control embryo with only an aluminum insert (red line), the notochord (green line) and ventral half of neural tube (blue line) have rescued the embryos from the deficiency at least when they have been
operated at SM-SP or SM stage, while the dorsal half of neural tube promoted the malformation (yellow line). while normal QT6 cells did not (Fig.4D; arrow head indicates missing proximal rib)), suggesting that proximal rib development depends on the notochord and the ventral neural tube, and this effect might be mediated by Shh secreted by these axial tissues. Role of the surface ectoderm in development of the distal part of the rib We have reported that blocking the interaction between the
somite and the somatopleure caused deficiency in the sternal rib
(Sudo et al., 2001). However, the vertebral ribs were intact. That is,
at least the proximal half of the distal rib development does not
depend on the somatopleure. We therefore examined another
tissue surrounding the somite, the surface ectoderm. Block of somite-ectoderm interaction Because aluminum foil is too thick to insert between surface
ectoderm and somatic mesoderm, we used polyethylene
terephthalate (PET) film as a barrier. A piece of PET film about
0.25-0.33 mm by 1 mm was inserted between the surface ectoderm
and the thoracic somitic mesoderm at SM-SP stage in two-day chick embryos (Fig.5A; between green arrows). The width of the
film was about twice the diameter of somite. Consequently, at the
time of operation the PET insert was large enough to cover the
somite. However, 24 hours after the operation, as the embryo
grew, the width of the film became equal to the somite diameter.
Besides, the film tended to move laterally. Therefore we checked
the position of the film 24 hours after the operation to know whether
the film covered the somite at least for the 24 hours.
Seven days after the operation, the distal ribs were found to be
deficient (Fig.5B; red arrowheads) in all the embryos keeping the
PET insert between the ectoderm and the somitic mesoderm for
the first 24 hours (n=13)(Table 1, case 1 + case 2 + case 3). Among
these, in three embryos not only the somitic mesoderm but also
part of the lateral plate mesoderm were covered (Table 1, case 2)
and in seven cases the somite and part of the neural tube were
covered (Table 1, case 3) with the insert for 24hours.
When the insert covered only part of the somite and lateral plate
for the first 24 hours, the ratio of embryos lacking the distal ribs to
total operated number were 6/ 21 (29%) (Table 1, case 4), while
covering only the lateral plate did not cause the loss of distal rib
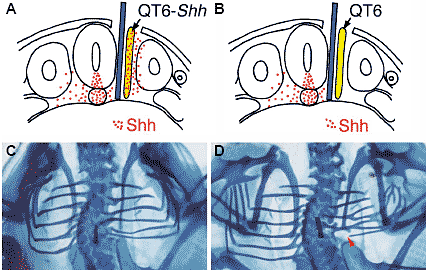 Fig. 4. Co-transplantation experiment showing
Shh can rescue the embryo from the proximal rib
deficiency caused by a blocked interaction
between the somitic mesoderm and the axial
tissues. (A) Schematic drawing of the transverse
section of a chick embryo operated at 2 days of
incubation. Red dots indicate Shh distribution. After
insertion of an aluminum foil barrier (blue bar) between
the axial tissues and the somitic mesoderm, Shhproducing
QT6 cells (QT6-Shh ; yellow bar with red
dots) were transplanted between the aluminum insert
and the somitic mesoderm. (B) Control operation.
Instead of QT6-Shh cells, normal QT6 cells were
transplanted. (C,D) Whole mount alcian blue staining
of the embryos 7 days after the operation. Dorsal
view. In the embryo with QT6- Shh cells, the ribs
connected to the vertebral column and were
considered to have the proximal rib (C), while the
embryo with normal QT6 cells (D) showed a deficient
proximal rib (red arrow head in D). Fig. 4. Co-transplantation experiment showing
Shh can rescue the embryo from the proximal rib
deficiency caused by a blocked interaction
between the somitic mesoderm and the axial
tissues. (A) Schematic drawing of the transverse
section of a chick embryo operated at 2 days of
incubation. Red dots indicate Shh distribution. After
insertion of an aluminum foil barrier (blue bar) between
the axial tissues and the somitic mesoderm, Shhproducing
QT6 cells (QT6-Shh ; yellow bar with red
dots) were transplanted between the aluminum insert
and the somitic mesoderm. (B) Control operation.
Instead of QT6-Shh cells, normal QT6 cells were
transplanted. (C,D) Whole mount alcian blue staining
of the embryos 7 days after the operation. Dorsal
view. In the embryo with QT6- Shh cells, the ribs
connected to the vertebral column and were
considered to have the proximal rib (C), while the
embryo with normal QT6 cells (D) showed a deficient
proximal rib (red arrow head in D). 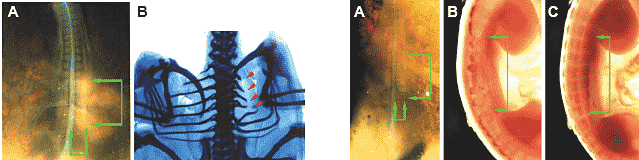 Fig. 5 (Left). Deficiency in the distal rib caused by blocking the interaction between the thoracic somitic mesoderm and the surface ectoderm in 2 day chick embryos. (A) Dorsal view of the embryo immediately after the operation. Upper is rostral. A piece of polyethylene terephthalate (PET) film (ca. 1/4-1/3 mm X 1 mm) was inserted underneath the surface ectoderm on the somitic mesoderm including both segmented somites and unsegmented segmental plate. PET film is indicated by two pairs of green arrows. (B) Dorsal view of the embryo 7 days after the operation. Whole mount alcian blue staining. On the operated site, only the proximal ribs were formed and the distal ribs were missing (red arrowheads). Fig. 6 (Right). Pax3 expression pattern in the embryo with blocked interaction between thoracic somitic mesoderm and surface ectoderm. (A) Dorsal view of the embryo immediately after the operation. Details are in the legend for Fig. 5A. (B,C) Whole mount in situ hybridization using Pax3 probe performed 2 days after the operation. Lateral side view. Upper is rostral. On the operated side (B), Pax3 expression is down regulated, compared with that on the control side (C). (n=14) (Table 1, case 5), except for only the loss of the sternal rib in three cases, as has been reported previously (Sudo, et al., 2001). In the case when the insert covered both the somite and the neural tube partially, 21% (4/19) of the operated embryos lacked their distal ribs (Table 1, case 6). Thus, the block of the interaction between the surface ectoderm
and the somitic mesoderm always resulted in the lack of the distal
rib, while the partial block of this interaction caused distal rib
deficiency with a much lower incidence. On the other hand, the
block of the interaction between the ectoderm and the lateral plate
or the neural tube was less effective in cases 4 and 6 than in cases
2 and 3. Since the inserts were approximately the same size, they
covered a wider range of tissues other than the somitic mesoderm
in cases 4, 6 than in cases 2, 3. These findings suggest that the
development of the distal rib requires the interaction between the
surface ectoderm and the somitic mesoderm. Pax 3 expression pattern
Two days after the operation, the expression pattern of Pax3,
characteristic in the dermomyotome, was examined by whole
mount in situ hybridization. Pax3 expression in the somites was
down regulated at the site where the PET film was inserted (Fig.6B;
between arrows), compared to the control side (Fig. 6C), suggesting
the block of somite-ectoderm interaction caused deficiency in the
dermomyotome. We have described elsewhere changes of
expression pattern of some genes in somites after the operation
(Hirao and Aoyama, 2004). Discussion Two developmental units for the rib: the proximal rib and the distal rib В этом исследовании мы показали, что проксимальная часть ребра развивается в зависимости от хорды и вентральной части нервной трубки, тогда как развитие дистальной части зависит от поверхностной эктодермы. В последнее время ребро рассматривается как состоящее из двух частей, проксимальной и дистальной. Первая является маленькой частью и состоит из головки, шейки, tuberculum и короткой проксимальной части тела, тогда как дистальная часть ребра составляет основную часть его тела.
Проксимальная часть ребра происходит из каудальной половины сомита, тогда как дистальная часть ребра происходит из обеих сомитных половинок после ре-сегментации, как это следует из наблюдений над эмбрионами людей (Sensenig, 1949) и экспериментов по трансплантации (Huang et al., 2000a; Aoyama & Asamoto, 2000). Проксимальная и дистальная часть ребра, как было показано, происходят из медиальной и латеральной половин сомита, в экспериментах с курица/перепел химерами или при DiI мечении (Olivera-Martinez et al., 2000). Хотя дистальная часть ребра может не происходить из собственно дермомиотома (Huang et al., 2000b; Evans, 2003), она появляется очень близко или на границе между дермомиотомом и склеротомом (Kato & Aoyama, 1998; our unpublished data), согласно нашим трансплантационным экспериментам.
Разные гены, как предполагается, контролируют независимо проксимальную и дистальную часть ребра. Используя Danforth's short-tail, мышей с мутантной и undulated хордой, Pax-1 мутантных мышей, Koseki et al. (1993) показали, что Pax1 отвечает за развитие проксимальной части ребер. С др. стороны, мутации в мышце-специфических транскрипционных факторах часто вызывают уродства в дистальных частях рёбер. В самом деле, Myf5 нокаутные мыши лишены дистальных частей ребер
TABLE 1 LACK OF THE DISTAL RIB CAUSED BY BLOCKING THE SURFACE ECTODERM-SOMITIC MESODERM INTERACTION 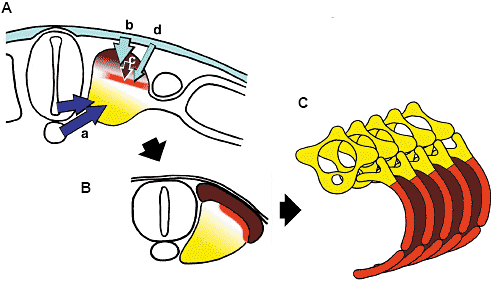 Fig. 7. The proximal rib and the distal rib development. Proximal rib development (yellow in ABC) depends on the notochord and/or
the ventral part of neural tube, presumably the floor plate. Distal rib
development (red in ABC) depends on the surface ectoderm. The surface
ectoderm may influence the distal rib rudiment directly (A, d) or indirectly through the dermomyotome (A, bc). The intercostal muscles are derived from the dermomyotome that arises from the dorsal part of each somite (brown in ABC). Fig. 7. The proximal rib and the distal rib development. Proximal rib development (yellow in ABC) depends on the notochord and/or
the ventral part of neural tube, presumably the floor plate. Distal rib
development (red in ABC) depends on the surface ectoderm. The surface
ectoderm may influence the distal rib rudiment directly (A, d) or indirectly through the dermomyotome (A, bc). The intercostal muscles are derived from the dermomyotome that arises from the dorsal part of each somite (brown in ABC). полностью (Braun et al., 1992). Осталось неясным, сам ли ген Myf5 отвечает за развитие ребер (Kaul et al., 2000); однако, по крайней мере, развитие дистальной части ребра контролируется геном(ами), который(ые) не отвечает за развитие проксимальной части ребра. Проксимальная и дистальная части ребер не только отличаются по их онтогенетическому происхождению и генам, необходимым для их нормального развития, но и по межтканевым взаимодействиям, необходимым для их развития. The neural tube and the notochord in the development of the
proximal rib Аксиальные ткани необходимы для развития осевого скелета (Teillet & Le Douarin, 1983; Teillet et al., 1998). SHH, продуцируемый хордой и донной пластинкой нервной трубки обеспечивает эти взаимодействия: Shh нулевые мутантные мыши обнаруживают тяжелые дефекты осевого скелета, не формируются компоненты ребер (Chiang et al., 1996). Депривация хорды нервной трубкой и Shh-нулевые нокауты дают очень сходные фенотипы. Эмбрионы восстанавливаются с помощью обратных трансплантаций SHH-продуцирующих QT6 клеток после удаления осевых тканей (Teillet et al., 1998). Т.о., SHH может замещать осевые ткани при формировании осевого скелета.
Т.к. в этом случае ребра полностью отсутствуют, если блокировать взаимодействие между осевыми тканями и сомитами, то затрагиваются только проксимальные части ребер, а SHH способен освобождать эмбрионы от этого недостатка. Это расхождение может быть обусловлено разными стадиями развития, на которых сомитная мезодерма лишается влияния аксиальных тканей. Когда мы изолировали сомитную мезодерму на стадии сегментной пластинки от аксиальных тканей, то мы часто обнаруживали полное отсутствие ребер (Fig 1D). Полное устранение аксиальных тканей у ранних эмбрионов, как было показано, ведет к гибели сомитных клеток (Teillet & Le Douarin, 1983), указывая тем самым, что ребра полностью искореняются, если эта депривация происходит до сегментации.
С др. стороны, если сомитная мезодерма изолируется от влияния аксиальных тканей после того, как началась сегментация (SM-SP, SM
стадия), то отсутствуют только проксимальные части ребер, как это имеет место у мутантных Pax1 мышей (Wallin et al., 1994). Pax1, как было показано, отвечает за формирование вентральной части осевого скелета, включая проксимальные части ребер (Wallin et al., 1994) и экспрессируется в сомитах после сегментации (Ebensperger et al.,
1995), среди субдоменов склеротома (Christ et al., 2004), поэтому только рудименты проксимальных частей ребер могут затрагиваться, если операция производится после сегментации. Huang et al. (2003) показали, что SHH косвенно регулирует развитие дистальных частей ребер посредством продуцируемого дермомиотомом FGF8. Такое влияние осевых тканей может происходить до дифференцировки дермомиотома, т.к. дистальные части ребер развиваются нормально, когда осевые ткани удаляются после сегментации на сомиты.
Т.о., наши находки подтверждают, что развитие проксимальных частей ребер зависит от хорды и/или вентральной части нервной трубки, эффект, который может обеспечиваться Shh, продуцируемого этими аксиальными тканями (Fig. 7A a). Surface ectoderm and development of the distal rib Удаление поверхностной эктодермы, как известно, вызывает разного типа уродства ребер (Huang et al., 2000b; Sudo et al., 2001). Ребра могут отсутствовать, укорачиваться или иметь неправильную форму, если поверхностная эктодерма, покрывающая торакальную сегментную пластинку, удалена (Huang et al., 2000b).
Huang et al. (2000b) полагают, что эти уродства ребер обусловлены отсутствием дермомиотома, т.к. развитие дермомиотома зависит от поверхностной эктодермы (Sosic et al., 1997; Dietrich et al., 1997; Hirao and Aoyama, 2004). Фактически, Huang et al. (2003) показали, что имплантации кусочков, пропитанных Fgf-8 усиливают образование реберных хрящей, тогда как SU 5402, ингибитор Fgf, обусловливает отсутствие реберных хрящей вблизи SU 5402 кусочков. Т.к. Fgf-8 экспрессируется в дермомиотоме (Huang et al., 2003), то отсутствие дистальных частей ребер, обусловленное устранением поверхностной эктодермы, может быть объяснено потерей латеральной части дермомиотома без влияния поверхностной эктодермы (Dietrich et al., 1997; Hirao and Aoyama, 2004).
Мы нашли, что дистальные части ребер всегда отсутствуют, если сомитная мезодерма изолирована от эктодермы с помощью PET пленки, по крайней мере, в течение 24 ч. Из-за роста эмбриона в течение этих 24 ч пленка д. покрывать не только сомитную мезодерму, но и также нервную трубку и/или латеральную пластинку мезодермы во время операции. Возникает вопрос, не может ли эктодерма влиять на развитие сомитов посредством нервной трубки и/или латеральной пластинки мезодермы. Однако, маловероятно, что ткани, соседствующие с сомитной мезодермой, играют важную роль во взаимодействии между эктодермой и сомитной мезодермой. Во-первых, значительно меньшее число эмбрионов лишено дистальных частей ребер в случае, более широкая область нервной трубки или латеральной пластинки покрывается пленкой, чем в случае, когда сомитная мезодерма покрывается пленкой в течение 24 ч (Table 1). Во-вторых, в данной работе изоляция сомитной мезодермы от нервной трубки обусловливала отсутствие только проксимальных частей ребер, а дистальные части оставались интактными, если операция осуществлялась на сомитной мезодерме на стадии SM-SP, подтверждая, что дистальные части ребер могут развиваться независимо от нервной трубки, В-третьих, в отсутствие мезодермы латеральной пластинки, позвонковые части ребер не затрагиваются, тогда как стернальные части ребер отсутствуют (Sudo et al., 2001). Альтернативным подразделением ребер является деление на позвоночную часть и на стернальную часть ребра. Тогда дистальная часть ребра может быть подразделена на vertebral часть и sternal часть. Развитие только стернальной части ребер зависит от латеральной пластинки мезодермы. Т.о., по крайней мере, vertebral часть дистальной порции ребер развивается независимо от латеральной пластинки мезодермы. Развитие латеральной пластинки мезодермы зависит от поверхностной эктодермы. Если поверхностная эктодерма удаляется, то клетки латеральной пластинки погибают в результате апоптоза (Sudo et al., 2001).
Т.о., дистальные порции ребер, особенно их vertebral конец, нуждаются во взаимодействии между поверхностной эктодермой и сомитной мезодермой для своего развития. Поверхностная эктодерма может контролировать развитие дистальных частей ребер косвенно путем регуляции дифференцировки дермомиотома (Fig. 7A, bc), хотя также возможно, что поверхностная эктодерма влияет на рудименты дистальных частей ребер непосредственно (Fig. 7A, d). 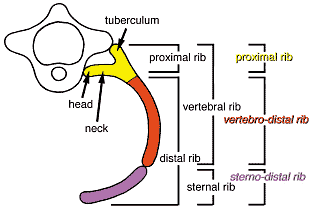 Fig. 8. Three compartments of the rib. The rib is considered to be composed of the proximal and the distal rib, or of the vertebral and the sternal rib. Combination of these two classifications makes three
compartments, which are shown to develop under different controls.
Proximal rib development depends on the ventral-axial tissues.
Development of the vertebral rib part of the distal rib, to which we refer
as vertebro-distal rib, depends on the surface ectoderm. Development of
the sternal rib part of the distal rib, to which we refer as sterno-distal rib, depends on the lateral plate mesoderm. Sterno-distal rib development
depends on the surface ectoderm indirectly, because differentiation and
survival of lateral plate mesoderm requires surface ectoderm. Fig. 8. Three compartments of the rib. The rib is considered to be composed of the proximal and the distal rib, or of the vertebral and the sternal rib. Combination of these two classifications makes three
compartments, which are shown to develop under different controls.
Proximal rib development depends on the ventral-axial tissues.
Development of the vertebral rib part of the distal rib, to which we refer
as vertebro-distal rib, depends on the surface ectoderm. Development of
the sternal rib part of the distal rib, to which we refer as sterno-distal rib, depends on the lateral plate mesoderm. Sterno-distal rib development
depends on the surface ectoderm indirectly, because differentiation and
survival of lateral plate mesoderm requires surface ectoderm. Three compartments of the rib: dependency on the ventroaxial
tissues, on the surface ectoderm and on the lateral plate
mesoderm Ранее мы сообщали, что развитие стернальных частей ребер зависит от латеральной пластинки мезодермы, а vertebral частей ребер нет (Sudo et al., 2001). Это подтверждает, вместе с представленными здесь данными, что ребра состоят из трех компартментов (Fig. 8), проксимальной части ребра, чьё развитие зависит от вентральных частей аксиальных тканей, vertebral части дистальной порции ребра (vertebro-distal rib ), чьё развитие зависит от поверхностной эктодермы, и стернальная часть ребра (sterno-distal rib ), чьё развитие зависит от латеральной пластинки мезодермы, которая, в свою очередь, зависит от поверхностной эктодермы. Эти три компартмента ребер совпадают с тремя онтогенетическими источниками, выявляемыми с помощью частичных удалений частей дермомиотома (Kato and Aoyama, 1998), хотя сам дермомиотом может и не давать зачатка ребра (Huang et al., 2000b). Region specific morphogenesis in the axial skeleton and
diversity of the axial skeleton: primaxial / abaxial skeleton Приняв, что ребро является онтогенетически сложным, состоящим из трех компартментов, может предположить, что длинная дистальная часть ребра, включающая стернальную порцию, может быть ответственна за регион-специфический морфогенез осевых скелетных единиц, которые, в свою очередь, ответственны за видо-специфические характеристики. Родоначальники млекопитающих и птиц могли потерять дистальные части ребер в области иной кроме торакса в ходе филогении.
Недавно, Burke and Nowicki (2003) предложили новую классификацию производных сомитов. Традиционно, мышцы туловища классифицировались на две категории, epaxial и hypaxial мышцы. Epaxial мышцы, свойственные мышцам спины, характеризуются своеи положением, дорсальнее по отношению к thoracolumbar fascia и своей иннервацией посредством дорсальных веточек спинальных нервов (Standring, 2005). Однако, недавние исследования с использованием мышей с мутантными производными сомитов показали несоответствие между фенотипами мутантов и epaxial/hypaxial классификацией. Напр., у Myf5 мутантных мышей затрагиваются и epaxial (vertebral) и hypaxial (intercostal) мышцы (Kablar et al., 1997). Поэтому Burke and Nowicki (2003) предложили новые категории, primaxial и abaxial.
Они определяют primaxial как сомитные клетки, которые развиваются исключительно в сомитных условиях, и abaxial как сомитные клетки, которые развиваются в производные латеральной пластинки мезодермы. Они классифицируют vertebral ребра с их межреберными мышцами как primaxial, а стернальные части ребер с их межреберными мышцами и абдоминальными мышцами, а также компонентами конечностей, как abaxial.
В соответствии с primaxial/abaxial классификацией может рассматривать, что у млекопитающих и птиц primaxial vertebro-distal части ребер уменьшаются в поясничной (abdominal) области, тогда как
abaxial sterno-distal части ребер с их ассоциированными мышцами могут меняться в абдоминальные мышцы и/или мышцы конечностей. Т.о., регион-специфичный, а также видо-специфичный морфогенез может рассматриваться как результат изменчивости каждого компартмента развития, primaxial/abaxial в стенке туловища, proximal rib|vertebro-distal rib|
sterno-distal rib.
|