вызывают молчание, тогда как одиночная копия нет (Stam et al., 2002), и сходным образом, почему два тандемных повтора необходимы для РНК-управляемого метилирования ДНК трансгена
(Chan et al., 2006). Robert Martienssen выдвинул гипотезу, почему тандемные повторы важны для поддержания молчания в центромерном гетерохроматине (Martienssen, 2003), которое может быть применено к
парамутациям. Множественные раунды RdRP и dicer-подобной активности с тандемными повторами в качестве матриц д. поддерживать увеличивающиеся пулы siRNA, считываемых со всей последовательности. Напротив с последовательности из одной копии последующие раунды амплификации будут продуцировать все более короткие и короткие dsRNAs. Др. модель заключается в том, что более крупные РНК, синтезируемые с повторов ответственны за молчание, которое не может быть генерировано с последовательности из одной копии. Третья идея заключается в том, что уникальные соединения фрагментов, сожщаваемые тандемными повторами, обладают специфическими свойствами, которые позволют осуществлять молчание.
Довольно неожиданно, почему B-I аллель, которые продуцирует siRNAs с его тандемных повторов, не заставляет молчать самого себя при высокой частоте. Хотя ничего, чтобы предупреждало замалчивание самого себя в B-I не было изучено — B-I является нестабильным, изменяясь в B' с частотой 0.1%-10% (Coe, 1966 and V.LC, unpublished data). Елдинственная возможность в том, что специфические белки могут активно предупреждать молчание. Напр., McGinnis et al.. (2006) неджавно показали, чтот состояние хроматина может становиться "immune" для молчания. Они описали две мутации, дефектные по парамутациям (включая
mop1-1), которые были способными реактивировать молчащий трансген, так что он становится транскрипционно активным в течение многих поколений, даже после того как белки дикого типа повторно вносили путем outcrosses. Др. возможрность в том. что тандемные повторы в B-I состоянии локализуются в разных ядерных компартментах, в которых аппарат молчания неспособен функционировать. Эти две возможности могут также объяснить отсутствие молчания в одиночной копии аллеля также. Недавнее исследование на
Arabidopsis показало, что ключевые компоненты путей siRNA модификаций хроматина коныцентрируются в специфических местах внутри ядра, которые могут быть факториями замалчивания (Li et al., 2006; Pontes et al., 2006).
Paramutation in Mice: Is RNA the Heritable Molecule?
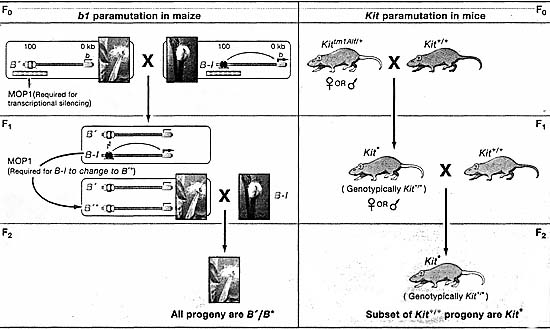
Недавно парамутация была описана у мышей в локусе Kit
(Rassoulzadegan et al., 2006). Локус Kit кодирует tyrosine kinase рецептор, который функционирует в мелоаногенезе, дифференцировке зародышевых клеток и гематопоэзе. В то время как мутантные мыши, лишенные Kit погибают в скоре после рождения, гетерозиготы с одним аллелем дикого типа и одним аллелем, не продуцирующим белка, имеют белый кончик хвоста и белые лапки (Figure 1B). Удивительно, когда гетерозиготы были скрещены др. с др. или мышами дикого типа, то большинство потомство генетически дикого типа имели белыми кончик хвоста и лапки и имели пониженные уронви мРНК Kit, сходные с теми, что у гетерозиготных родителей. Потомство с этим paramutant фенотипом было обозначено Kit*.
Здесь также передача парамутантного состояния, как полагают, связана с РНК. Однако, её роль у мышей, по-видимому, очень отличается от того, как она функционирует в локусе b1 растений. Rassoulzadegan et al. (2006) предположили, что молекулы РНК передаются посредством гамет и что они запускают деградацию Kit мРНК у индивидов с парамутацией. Как в парамутантном Kit*, так и гетерозиготном потомстве обнаруживается двухкратное снижение Kit мРНК отнросительно дикого типа. Авт. также налюдали накопление не полиаденилированных молекул РНК аномального размера у гетерозиготных Kit мышей. В спермиях гетерозиготных самцов имеется значительно больше Kit мРНК, чем обнаруживается ей у гомозиготных самцов дикого типа (сходные эксперименты не были описаны для Kit*). Чтобы протестировать, может ли увеличение Kit РНК отвечать за индукцию парамутантного фенотипа, исследователи инъецировали РНК от индивидов или гетерозиготных или гомозиготных по Kit дикого типа в одноклеточные эмбрионы дикого типа. Из тех, что дожили до рождения фенотип "white tip" чаще наблюдался у эмбрионов,которым инъецировали РНК от гетерозигот. Хотя фенотип белый кончик наблюдался в меньшей степени и при контрольных инъекциях РНК от Kit мышей дикого типа, фенотип редко передщавался потомству. С момощью инъекций microRNAs (miRNAs), которые деградируют Kit мРНК, они оказалось возможным генерировать white tip фенотип с частотами более высокими, чем инъекции неспецифических контрольных miRNAs. Способность продуцировать white tip фенотип с помощью микроинъекций РНК и miRNA в эмбрионы, позволила авт. постулировать, что эпигенетическое наследование ассоциирует с зиготическим переносом молекул РНК.
Другие два описанных случая парамутация-подобного наследования у мышей также были аллель специфичными и ассоциировали с конструированными аллелями (Rassoulzadegan et al., 2002; Herman et al., 2003; reviewed in Chandler and Stam, 2004). Сконструированный Kit аллель, изученный Rassoulzadegan et al. (2006) продуцировал уникальную мРНК с β-galactosidase кодирующей областью под контролем промотора Kit и регуляторной последовательности. Авт. сообщили, что др. сконструированный локус (в данном случае, содержащий GFP-neo cassette в первом интроне гена Kit) также генерировал парамутантное потомство, тогда как классическая точковая мутация в Kit нет. Означает ли это, что сконструированный Kit аллель, но не точковая мутация, также вызывает избыточную продукцию аберрантной РНК? Дальнейшее сравнеие транскрипции и структуры ДНК аллелей, которые индуцируют парамутации, с теми, которые обнаруживают Менделевское наследование, д. помочь даленйшему тестированию гипотезы.
Остается открытым вопрос, отличается ли Kit система у мышей фундаментально иным механизмом продукции парамутаций, чем у кукурузы с генным молчанием, происходящим на посттранскрипционном уровне скорее, чем на транскрипционном. Сходным образом остается открытым вопрос, какого типа молекулы обеспечивают наследуемость. В B' системе имеются четкие различия в хроматине между B' и B-I и это создает прецендент для РНК-обусловленных изменений в хроматине, нет нужды привлекать РНК в качестве молекулы наследственности. Однако, нет экспериментов на кукурузе, которые исключили бы передачу РНК. Для Kit системы разумно предположить, что перенос РНК с гаметами самцов и самок ведет к деградации транскриптов дикого типа. Однако, имеющиеся данные согласуются также с моделью, согласно которой инъецированная РНК (или РНК, продуцируемая с сконструированного аллеля) produced from the engineered allele) создают такое состояние хроматина, которое приводит к редукции транскрипции у Kit* мышей. Посттранкрипционное по сравнению транскрипционным молчанием д.быть отличимо с помощью ядерного run-on assays на мышах Kit*.
Даже если miRNA обладает способностью деградировать Kit мРНК, неясно, как miRNAs обеспечивают парамутации в сконструированных Kit аллелях: существуют множественные пути для достижения двукратного снижения Kit мРНК в потомстве white tip фенотипом. Авт. исслдедовали модификации хроматина, но не обнаружили заметных различий в метилировании ДНК или модификациях гистонов между диким типом и гетерозиготами и парамутантными животными внутри минимального Kit промотора. Однако, т.к. они не исследовали какие-либо др. области внутри и вокруг гена Kit, то они отмечали, что роль хроматина исключить нельзя.
Why Does Paramutation Exist and Is It Rare?
Возможно, что парамутации представляют собой редкие случаи, при которых нормальный регуляторный процесс, как тот, что регулирует центромерный и др. гетерохроматины, или пути защиты, призванные регулировать вирусы, действуют благодаря ошибке на редкие эухроматиновые гены или трансгены, содержащие чужеродную ДНК. Принимая во внимание превалирование siRNA путей (see M. Zaratiegui et al.. page 763 of this issue), почему парамутации не обнаруживаются в большем числе локусов? Эндогенный FWA локус может принимать одно из двух стабильных эпигенетических состояний: или тандемные повторы в промоторе метилируются и ген вынужден молчать или повторы неметилируются и ген активен. Системы b1 и FWA сходны наличем тандемных посторов, которые транскрибируются и генерируют siRNAs, кроме того, метилированный, молчащий FWA аллель не парамутирует неметилированный активный аллель; гетерозиготы передают два аллеля неизменными. Это так несмотря на тот факт, что когда вносится неметилированный трансген с тандемными повторами, то он становится метилированным. Как "естественный" активный аллель защищен от такого молчания, к которому чувствителен трансген? Защищено ли большинство генов от такого типа молчания? B-I может предсавлять крайний пример аллеля, который, сильно сенсибилизирован, чтобы стать молчащим, т.к. он ошибочно выглядит как чужеродная ДНК, с помощью защитной системы клеток.
Возможно также, что парамутации возникают чаще, чем предполагается. Признаком нескольких систем парамутаций является то, что лекие отклонения в уровнях экспрессии лего выявляются с помощью видимых маркеров, таких как регуляторные гены, затрагивающие пигментацию у растений или окраску шерсти у мышей. Необходимо также подчеркнуть, что чрезвычайная пенетрантность и наследуемость B-I, которые невозможно игнорировать, являются необычными случаями (rev. Chandler and Stam, 2004). Большинство др. парамутаций, как это наблюдается с Kit, были фенотипически не полностью пенетрантны и терялись после нескольких поколений случайных скрещиваний. Низкая пентрантность и нестабильность сконструированных Kit аллелей, изученных Rassoulzadegan et al. (2006) может объяснить, почему полученные более декады тому назад, они свою способность индуцировать не-Менделевское наследование, проявили только недавно. Итак, комбинация немногих мощных генетических маркеров, изменчивость и нестабильность событий и наблюдения, что парамутации появляются только в специфических аллелях, всё это может вносить вклад в неспособдность распознавать др. примеры. Использование геномных маркеров и проверка паттернов аллель-специфичной экспрессии в ряде поколений может служить одним из подходов для поиска парамутаций на шкале всего генома.
Возникают или нет парамутации благодаря клеточному механизму защиты. Авт. склоняется к тому, что парамутации сегодня представляют собой фундаментальный механизм регеуляции гнов и наследственности. Имеется несколько потенциальных ролей для аллель- или гомолог-зависимого переноста эпигенетической информации потомству: он может представлять адаптивный механизм для переноса благоприятных (индуцированных внешней средой?) состояний экспрессии потомству или может использоваться для установления функциональной гомозиготности у полиплоидов и может частично объъяснить снижение приспособленности, связанное с инбридингом (rev. Chandler and Stam, 2004). Сходный с парамутациями феномен также может вносить вклад в низкую пенетрантность не-Менделевского наследования, часто ассоциированного со сложными болезнями у человека.
В ближайшее время мы не только достигнем лучшего механистического понимания парамутаций в современных системах, но и столкнемся с ноткрытием многих новых примеров эпигенетических механизмов передачи информации, которая регулирует генную экспрессию в ряде поколений.
Сайт создан в системе
uCoz