Термин "self-tolerance" обозначает все механизмы, которые защищают тело от атак с помощью свой собственно иммунной системы. Адаптивный отдел иммунной системы генерирует иммунные клетки, которые экспрессируют антиген-специфичные рецепторы за счет случайного механизма, который нуждается в контроле качества - отборе "personalized" репертуара рецепторов, направленных против чужеродных, но не против собственных антигенов (1). Центральная и периферическая толерантность к себе отличаются в соответствии с местом, которого касается толерантность (2). Центральная толерантность для Т лимфоцитов осуществляется в тимусе, где генерируется их первичный репертуар рецепторов антигенов. Здесь развивающиеся Т клетки, которые распознают и и реагируют на собственные антигены элиминируются или превращаются в Т регуляторные клетки, которые супрессируют активацию иммунной системы и предупреждают само-реактивность. Хотя тимус обладает обширным запасом само-антигенов, включая и те, чья экспрессия иным образом ограничивается специфической тканью, эта коллекция тем не менее неполная. В этом номере, Gardner et al. (3) описывают, как периферическая лимфоидная ткань действует как сеть безопасности, предупреждая Т клетки, специфичные по антигенам, не присутствующим в тимусе, от возможности избежать элиминации.
Медуллярные тимические эпителиальные клетки, особенно тип стромальных клеток тимуса, экспрессируют разнообразные наборы генов, которые каким-то образом ограничены определенной тканью и/или стадией развития (4). Эта т.наз. разнородная генная экспрессия в тимусе частично регулируется с помощью транскрипционного регулятора, наз. Autoimmune regulator (Aire). Мыши, дефицитные по Aire, характеризуются мультиорганным аутоиммунным синдромом, сходным с таковым у людей с функциональными мутациями в гене Aire gene (5).
Aire экспрессируется на высоком уровне в медуллярных эпителиальных клетках тимуса. Однако функция соотв. экспрессии
Aire в клетках периферических лимфоидных органов спорна (5-9). Gardner et al. идентифицировали клетки периферических лимфатических узлов, селезенки и Пейеровых бляшек (лимфоидных структур кишечника), которые экспрессируют
Aire и обеспечивают делецию аутореактивных Т клеток. Авт. генетически преобразовали мышей, у которых промотор гена
Aire управляет экспрессией слитого белка, состоящего из green fluorescent protein и islet-specific glucose-6-phosphatase related protein (Igrp), антигена, специфичного для поджелудочной железы. Из медуллярных эпителиальных клеток тимуса и периферических клеток, которые экспрессируют репортерный белок, 85% и 25% экспрессируют эндогенные
Aire, соотв. Большинство из этих периферических клеток, называемых внетимусными
Aire- экспрессирующими клетками, были эпителиальными клетками стромального типа, расположенными в интерфейсе между областями T и B клеток в периферических лимфоидных тканях. Эти клетки экспрессируют также рецепторы, характерные для антиген-презентирующих клеток, но отличаются по некоторым маркерам от медуллярных эпителиальных клеток тимуса. Неожиданно некоторые из таких внетимусных Aire-экспрессирующих клеток были высоко мобильны внутри микросреды лимфатических узлов и в то же самое время были способны делетировать Т клетки, специфические по репортерному белку. Вообще-то наиболее интригующим результатом этого исследования стало то, что гены мишени, контролируемые с помощью Aire, в тимусные, чем на периферии. Количество генов в последней примерно 1/10 от того, что в тимусе и их степень Aire-зависимой регуляции менее выражена. Более того, хотя и имеется мало перекрывания между генными пулами, оба пула безусловно обогащены генами, кодируемыми для тканеспецифических само-антигенов (see the figure). Характерный состав обоих генных пулов говорит в пользу роли периферической толерантности, которая
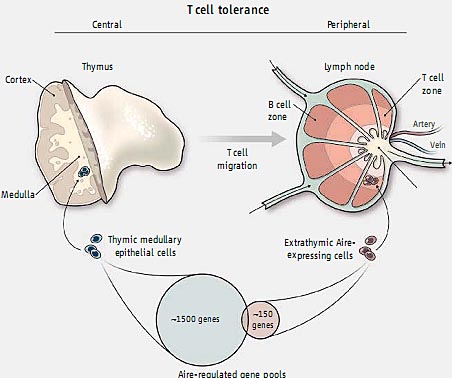 Complementary tolerance. The transcriptional regulator Aire controls the expression of complementary pools of self-antigens in the thymus and peripheral lymphoid tissues that sequentially imprint central andperipheral T cell tolerance, respectively.
Complementary tolerance. The transcriptional regulator Aire controls the expression of complementary pools of self-antigens in the thymus and peripheral lymphoid tissues that sequentially imprint central andperipheral T cell tolerance, respectively.
дополняет толерантность, развиваемую в тимусе. Недавнее исследование Lee et al. also также идентифицировало фракцию не-гематопоэтических клеток в мезентерических лимфатических узлах, которые экспрессируют
Aire и определенные тканеспецифические само-антигены и обеспечивают делецию периферических Т клеток (9). Однако эти клетки были менее сильно обогащены и дифференцировались фенотипически в те, что сравнимы с идентифицированными Gardner et al. Более того, данные Lee et al. находятся в большом соответствии с концепцией, что периферическая толерантность служит как резерв центральной толерантности скорее, чем оказывается дополнительной.
Исследование Gardner et al. всё ещё оставляет некоторые нерешенные вопросы. Почему только 25% периферических клеток у трансгенных мышей, которые экспрессируют флюоресцентный репортерный белок, экспрессируют также эндогенный Aire? Обусловлено ли это эктопической экспрессией репортерной конструкции в общем-то Aire-негативных клетках? Является ли эндогенная экспрессия Aire слишком низкой, чтобы быть выявляемой или экспрессия Aire и репортерного белка не синхронизированы? Относительно низкое совпадение между экспрессией репортера и Aire может также вносить вклад в кажущуюся довольно низкой степень генной экспрессии, индуцируемой Aire в периферических клетках, которая для большинства генов меньше, чем вдвое по сравнению с фоновой экспрессией у Aire-дефицитных мышей.
Хотя индукция толерантности, как полагают, чрезвычайно чувствительна к низким количествам само-антигенов, которые представлены Т клеткам в контексте основного комплекса гистосовместимости, очень важно показать, что низкие уровни экспрессии эндогенных ткане-специфических само-антигенов в периферических Aire-экспрессирующих клетках являются "tolerogenic."
Итак, на первый взгдяд, аутоиммунный фенотип у Aire-дефицитных мышей полностью воспроизводится с помощью трансплантации Aire-дефицитных стромальных клеток тимуса (при отсутствии функциональных доказательств Aire внетимусных сайтов) (5). Учитывая разный состав само-антигенов, характеризующих периферические клетки, аутоиммунный фенотип, вызываемый отсутствием Aire на периферию, может быть едва заметным и, как правило, не выявляется. Несмотря на это имеются предостережения, исследование Gardner et al. ставит интригующие вопросы о роли и функции Aire и природе периферических клеток, которые экспрессируют этот фактор (10).
Появление внетимусных Aire-экспрессирующих клеток у позвоночных интересно, учитывая, что организованный вторичные лимфоидные органы участвуют значительно позднее, чем тимус (11). Aire представлен одиночной копией гена с ортологами у млекопитающих, птиц и рыб, чья структура законсервирована у позвоночных свыше 400 миллионов лет (12). Не описано родоначальных Aire генов у беспозвоночных. Это указывает на то, что Aire и его роль в толерантности приобретены рано во время эволюции позвоночных, скорее всего в конкуренции с появлением адаптивной иммунной системы. Возникает вопрос, является ли роль Aire's лишь гарантией центральной толерантности или она может быть кооптирована с др. функциями. Работа Gardner et al. предоставляет строгий аргумент в пользу последнего -Aire также, по-видимому, вносит вклад в установление периферической толерантности.
Сайт создан в системе
uCoz 