Недавние исследования показали, что коричневый жир и мышцы могут иметь много общего. У млекопитающих белая жировая ткань хранит энергию в форме жира и это связано с ожирением, тогда как коричневый вариант сжигает жир, чтобы генерировать тепло и тем самым противодействует ожирению. Стимуляция увеличения коричневого жира у людей может, следовательно, служить потенциальным средством лечения ожирения, но сначала необходимо до конца понять онтогенетическое происхождение двух типов жировой ткани.
Seale et al. использовали короткую hairpin РНК в клетках предшественниках, чтобы послать в нокдаун , транскрипционный регулятор коричневых жировых клеток, и наблюдали, что маркеры клеток коричневого жира репрессированы, тогда как миогенные гены активировались. Более того, клетки превращались в bona fide мышечные клетки с морфологией слитых мышечных трубок. Интересно, что предприняли эксперименты по отслеживанию клонов у мышей, которые были генетически модифицированы, чтобы обнаружить экспрессию myogenic factor-5 (Myf5), гена, специфичного для скелетных мышц. Клетки, экспрессирующие Myf5, формировали клетки скелетных мышц и коричневого, но не белого жира. Это указывает на то, что клетки коричневого жира возникают из предшественников скелетных мышц.
Форсированная экспрессия PRDM16 в клетках мышечных предшественников блокировала миогенез и индуцировала адипогенез коричевого жира. Чтобы исследовать механизм действия PRDM16, Seale et al. очищали PRDM16 белковый комплекс из жировых клеток и анализировали с помощью масс-спектрометрии. Они идентифицировали peroxisome proliferator-activated receptor-gamma (PPARγ), важный регулятор адипогенной дифференцировки в качестве партнера по связыванию с PRDM16. PRDM16 усиливал экспрессию PPARγ репортерного гена и был неспособен обеспечивать адипогенез в PPARγ-дефицитных фибробластах. Итак, PRDM16 стимулирует brown adipogenesis путем индукции PPARγ.
В независмом исследовании, Tseng et al. обрабатывали предшественников жировых клеток bone morphogenetic proteins (BMPs), которые важны для многочисленных процессов во время эмбрионального развития. Они установили, что один BMP7 может инструктировать предшественников клеток коричневого жира становиться зрелыми клетками, которые экспрессируют маркеры клеток коричневого жира и гены. которые участвуют в биогенезе и функции митохондрий. Интересно, что Prdm16 находился среди генов, активируемых с помощью BMP7.
BMP7 активирует сигнальный путь p38 mitogen-activated protein kinase (MAPK) в клетках предшественниках коричневого жира, а специфические ингибиторы этого пути блокируют индуцируемую с помощью BMP7 экспрессию маркеров клеток коричневого жира. Более того, BMP7-дефицитные эмбрионы и новорожденные мышей имеют редуцированную ткань коричневого жира, хотя др. органы были нормальными. Сходно с, Seale et al. наблюдалась аномальная морфология клеток коричневого жира и активация мышце-специфических генов у эмбрионов, которые были лишены PRDM16. При доставке с помощью вирусов BMP7 мышам дикого типа, Tseng et al. индуцировали увеличение массы коричневого жира, что приводило к более высоким энергетическим тратам и к уменьшению избытка веса по сравнению с контрольными животными.
Эти исследования указывают на то. что обработка людей BMP7 или увеличение уровней PRDM16, которые активируют дифференцировку коричневого жира и увеличивают энергетические траты, может быть новым способом борьбы с ожирением.
How Now, Brown Fat?
Mitchell A. Lazar ( lazar@ mail.med.upenn.edu) SCIENCE, 22 AUGUST 2008 VOL 321, No , P.1048 www.sciencemag.org 1048
Brown adipose tissue is, surprisingly, more related to skeletal muscle than to white adipose tissue.
Obesity, the condition of excess fat storage in adipose tissue, is harmful to health and epidemic in modern society (1). Adipocytes, or “fat cells,” are spe- cialized cells that store this fat in triglyceride- containing droplets. Until recently, it was thought that all adipocytes are derived from a common precursor. It seemed almost obvious that different types of fat cells are closely related. However, this week, a study by Seale et al. (2) provides compelling evidence against this doctrine.
Not all adipocytes have the same physiological function. Those in white adipose tissue constitute the major energy storage depot in mammals. When nutritional sources of energy are scarce, triglyceride breakdown in white adipocytes generates fatty acids that are exported and used as fuel by other tissues, including muscle. In obesity, where energy expenditure is matched, and usually exceeded, by caloric consumption, both the size and the number of white adipocytes increase. White adipose tissue is often classified on the basis of its location as subcutaneous and abdominal (or visceral) fat. Secretions from visceral fat pass through the liver before entering the gen- eral circulation, and this has been implicated in the pathogenesis of type 2 diabetes and insulin resistance (3). Visceral adipocytes dif- fer from subcutaneous adipocytes in their gene expression profiles (4), although there is no evidence that the precursors of white adi- pocytes in visceral and subcutaneous depots are fundamentally different.
In contrast to white adipose tissue, brown adipose tissue plays an active role in energy expenditure, oxidizing fatty acids produced by triglyceride hydrolysis to generate heat. As
the name suggests, brown adipose tissue appears different from white adipose tissue, largely because of the increased number of mitochondria (cytochromes within the mito- chondria account for the color) that function in oxidative metabolism. Also, brown adi- pocytes contain multiple small lipid droplets, whereas white adipocytes usually have a single large lipid droplet. The thermogenic oxi- dation of fatty acids is facilitated by uncou- pling protein–1, which is expressed specifi- cally in brown adipocytes. In rodents, brown fat is characteristically found in the inter- scapular region, and is critical for thermoreg- ulation throughout life. By contrast, humans have proportionally more brown fat at birth than in adulthood, when it can be difficult to locate. Nevertheless, recent studies point to the existence and conserved function of brown adipose tissue in humans (5). Although brown and white adipocytes look different and have different physiological functions, they are considered to have derived from a com- mon precursor cell. One rea- son is that the transcription factor peroxisome proliferator- activated receptorγ (PPARγ), which is the master regulator of adipogenesis, is required for differentiation of both white and brown adipocytes (6). In addition, conversion of white to brown adipocytes is induced by β-adrenergic stimulation— that is, stimulation of white adipose with hormones (cate- cholamines) that are released in situations of stress or low blood sugar concentration (7).
Paths to muscle and fat. Skeletal myocytes and brown adipocytes derive from a common precursor cell that expresses the transcription factor Myf5. White adipocytes derive from a Myf5-negative precursor, as do brown adipocyte–like cells that appear in white fat depots after adrenergic stimulation. These distinct cell types play very different roles in physiology.
The factors that mediate conversion of white to brown adipose tissue have been ex- tensively investigated, moti- vated, in part, by the hope that expansion of the brown-fat compartment, leading to in- creased energy expenditure, would be a novel way to treat obesity (8). Ten years ago, PPARγ coactivator 1α (PGC- 1α) was identified as a tran- scriptional coactivator that is enriched in brown adipocytes (9). Since then, PGC-1a has emerged as a major metabolic regulator in brown fat, as well as other tissues, although it is not required for the expression of other brown fat–specific genes, including uncoupling pro- tein–1 (10). About a year ago, in a more sys- tematic search for brown-fat transcription fac- tors, Seale et al. found that a previously unsus- pected protein, PRDM16, is required to main- tain the brown-fat phenotype, including the expression of uncoupling protein–1 (11).
The implication that PRDM16 and PGC- 1aare involved in the generation and function of brown adipocytes advanced the field, but did not alter the basic notion that brown and white adipocytes derive from a common pre- cursor. Therefore, the new finding from Seale and colleagues is a real surprise: Brown adipocytes actually derive from a cell that otherwise gives rise to skeletal muscle but not to white adipocytes (2). This conclusion is sup- ported by evidence that forced expression of PRDM16 leads cultured mouse skeletal mus- cle cells to differentiate into brown adipocytes and, conversely, that brown adipocytes de- pleted of PRDM16 take on the look and func- tion of skeletal muscle cells. Independent of genetic manipulation of PRDM16, lineage- tracing studies indicate that brown adipocytes are derived from cells that express the tran- scription factor Myf5, a molecular marker of skeletal muscle precursors. Importantly, white adipocytes did not derive from these cells, demonstrating that the developmental path- ways of white and brown adipocytes diverge before the switch between brown fat and skeletal muscle (see the figure).
The link between brown fat and skeletal muscle was presaged by the observation that gene-expres- sion profiles of brown adipose tissue share features characteris- tic of skeletal muscle (12). Indeed, in retrospect, the oxida- tive metabolism of brown fat, as well as its color and mitochon- drial content, is more similar to those of skeletal muscle than to those of white adipose tissue. The relationship between brown adipose tissue and skeletal mus- cle is likely to extend to humans, where brown adipocyte precur- sors have recently been identi- fied in skeletal muscle (13).
The close connection be- tween brown adipose tissue and skeletal muscle raises several questions. How does adrenergic stimulation convert white to brown adipose tissue? Seale et al. report that these brown adipocyte–like cells, although they express PRDM16, are not de- rived from Myf5-expressing precursors. Hence, there must be an alternative pathway to brown adipocyte formation that is more highly related to that of white adipogenesis. Are the brown adipocyte–like cells that emerge in white fat depots dis- tinguishable, molecularly or functionally, from the adipo- cytes in brown fat depots? Will understanding the origin of brown adipocytes lead to novel therapeutic approaches to obe- sity? Confronted with a seem- ingly inexorable rise in obesity and its dire medical consequences, our society may well benefit from new developments.
References
1. M. A. Lazar, Science 307, 373 (2005).
2. P. Seale et al., Nature, 454, 961 (2008).
3. A. H. Kissebah, G. R. Krakower, Physiol. Rev. 74, 761 (1994).
4. S. Gesta, Y. H. Tseng, C. R. Kahn, Cell 131, 242 (2007).
5. J. Nedergaard, T. Bengtsson, B. Cannon, Am. J. Physiol. Endocrinol. Metab. 293, E444 (2007).
6. M. Lehrke, M. A. Lazar, Cell 123, 993 (2005).
7. S. R. Farmer, Genes Dev. 22, 1269 (2008).
8. C. Tiraby, D. Langin, Trends Endocrinol. Metab. 14, 439 (2003).
9. P. Puigserver et al., Cell 92, 829 (1998).
10. B. N. Finck, D. P. Kelly, J. Clin. Invest. 116, 615 (2006).
11. P. Seale et al., Cell Metab. 6, 38 (2007).
12. J. A. Timmons et al., Proc. Natl. Acad. Sci. U.S.A. 104, 4401 (2007).
13. M. Crisan et al., Stem Cells 10.1634/stemcells.2008- 0325 (2008).
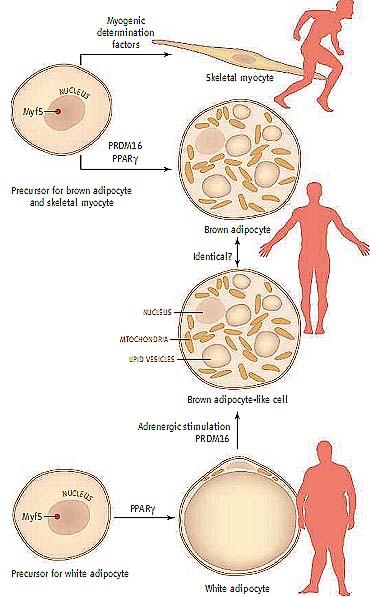 Paths to muscle and fat. Skeletal myocytes and brown adipocytes derive from a common precursor cell that expresses the transcription factor Myf5. White adipocytes derive from a Myf5-negative precursor, as do brown adipocyte–like cells that appear in white fat depots after adrenergic stimulation. These distinct cell types play very different roles in physiology.
Paths to muscle and fat. Skeletal myocytes and brown adipocytes derive from a common precursor cell that expresses the transcription factor Myf5. White adipocytes derive from a Myf5-negative precursor, as do brown adipocyte–like cells that appear in white fat depots after adrenergic stimulation. These distinct cell types play very different roles in physiology. 
