Enhancing Gene Regulation
 SCIENCE 5 SEPTEMBER 20O8, VOL 321, No 5894, P. 1300-1301 | |
|
Bioinformatic approaches reveal functional changes and the evolution of regulatory sequences that control gene expression.
|
В 1975 in an influential paper by Mary-Сlaire King and Alan Wilson (2), предположили, что различия в признаках между людьми и шимпанзе в первую очередь обусловлены регуляторными различиями в экспрессии генов. В данном номере Prabhakar et al. на стр. 1346 (3), и Hong et al. на стр. 1314 (4), продемонстрировли мощь комбинированных биоинформационных подходов с экспериментальными тестами, чтобы охарактеризовать регуляторные регионы.
Последовательности ДНК, которые регулируют транскрипцию генов, занимают не фиксированную позицию относительно кодирующих регионов ДНК и часто диффузны и широко разбросаны. Даже если позиция регуляторного элемента известна, то существуют затруднения идентификации. какие мутации вызывают функциональные последствия. В кодирующих последовательностях, генетический код накладывает хорошо знакомые закономерности: мутации, которые изменяют структуру белка могут быть идентифицированы исчерпывающе и однозначно. Напротив, идентификация мутаций внутри регуляторных областей требует экспериментальных тестов предполагаемых регуляторных элементов от разных видов или индивидов — дорогой и требующий затрат времени - затратный процесс. Биоинформационные методы открывают способ идентификации многообещающих функциональных некодирующих регионов и сужают фокус экспериментального тестирования.
Одним из подходов является исследование генома по высоко законсервированным блокам некодирующих последовательностей (исходя из предположения, что консервация подразумевает функцию) и последующим сканированием примеров быстрой дивергенции последовательностей непосредственно в одной филогенетической ветви (это указывает на функциональные изменения у одиночных видов) (5). Prabhakar et al. использовлаи этот подход для идентификации некодирующей области, которую они назвали human-accelerated conserved noncoding sequence 1 (HACNS1) (see the figure). Чтобы протестировать функцию этой области они содали мышиных эмбрионов, которые экспрессируют конструкцию, состоящую из HACNS1 человека, промоторного элемента хитшокового гена и репортерного гена. Они установили, что HACNS1 человека управляет экспрессией в мезенхиме ранней развивающейся передней конечности и позднее в развивающейся задней конечности мышиных эмбрионов. Сравнение управления паттернов экспрессии у макак, шимпанзе и людей ортологов HACNS1 показало, что соотв. строгая экспрессиия в передних конечностях является уникальным признаком человеческой версии. Путем тестирования различных комбинаций HACNS1 последовательностей шимпанзе и человека
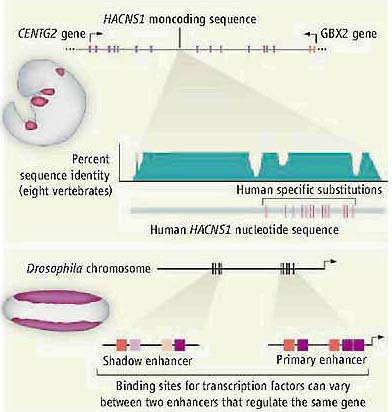 Identification of enhancer elements. (Top) HACNS1 is a noncoding region ol conservation (percent identity) among eight vertebrale species, with 13 human-specilic substitutions {vertical red lines). HACNSI drives the expression ol a reporter gene (purple) in the limbs of a developing mouse embryo. (Bottom) Primary enhancers (red and purple boxes) near a gene are conserved relative lo more distant "shadow" enhancers, which appear to be less functionally constrained (pink and light purple boxes). These enhancers drive gene expression (purple) in the Drosophila embryo.
Identification of enhancer elements. (Top) HACNS1 is a noncoding region ol conservation (percent identity) among eight vertebrale species, with 13 human-specilic substitutions {vertical red lines). HACNSI drives the expression ol a reporter gene (purple) in the limbs of a developing mouse embryo. (Bottom) Primary enhancers (red and purple boxes) near a gene are conserved relative lo more distant "shadow" enhancers, which appear to be less functionally constrained (pink and light purple boxes). These enhancers drive gene expression (purple) in the Drosophila embryo. авт. сузили исследуемые функциональные мутации до области в 81-пары оснований, содержащие 13 замен, которые возникли в ходе эволюции человека. Такая концентрация замен очень необычна по отношению генома в целом, она указывает на позитивную селекцию этой области во время происхождения человека. Эта консервативная область расположена внутри интрона гена CENTG2, который кодирует guanosine triphosphatase activating белок, который регулирует эндосомы (с мембраной связанные пузырьки, которые транспортируют материалы в клетке). Он также находится ~300 kb ниже следующего ближайшего гена, GBX2, который кодирует транскрипционный фактор, который экспрессируется в др. местах развивающихся конечностей. Если GBX2 в самом деле является мишенью для регуляции с помощью HACNS1, то его роль может оказатья захватывающей. Благодаря роль GBX2's в развитии конечностей авт. отмечают, что изменения в его экспрессии д.приводить к изменению анатомии конечностей у людей — продуцируя. напр., специализации рук, которые облегчают использование инструментов или к модификациям стоп, связанными с прямохождением.
Др. биоинформационный подход для идентификации регуляторных элементов - это поиск кластеров сайтов связывания для потенциальных транскрипционных факторов (6). Hong et al. исследовали данные иммунопреципитации хроматина, комбинированные с технологией микромассивов — так наз ChlP-chip анализом — генома плодовых мух (Drosophila melanogaster) в отношении мишеней регуляции сж
транскрипционного фактора Dorsal и известных кофакторов (4). Неожиданно они нашли, что многие гены мишени для Dorsal содержат не один, а два кластера сайтов связывания транскрипционного фактора, указывающие на присутствие множественных регуляторных областей со сходной функцией (see the figure). Хотя большинство экспериментально верифицированных энхансеров расположены внутри нескольких т.п.н. от гена, который они регулируют, некоторые из вторичных энхансеров обнаруживают тенденцию располагаться на расстоянии в десятки kilobases. Авт. использовали трансгенных мух, чтобы показать, что как ближайший, так и удаленный кластеры из связывающих сайтов управляют сходными паттернами экспрессии репортерных генов для двух генов, brk и sog.
Эволюционная роль этих "теневых энхансеров" неясна. Одна из возможностей, предположенных Hong et al., заключается в том, что они предоставляют возможность естественному отбору поправлять регуляторные последовательности, тогда как первичные энхансеры поддерживают существенную функцию гена. В согласии с этой гипотезой то, что последовательности "теневых энхансеров" эволюционируют более быстро, чем первичные энхансеры, это указывает на то, что они оперируют в условиях меньших функциональных ограничений. Интригующая возможность заключается в том, что "теневые энхансеры" хуже обеспечивают различия в экспрессии среди видов, и это можно проверить с помощью сравнительного экспериментального подхода, который Prabhakar с коллегами применили к HACNS1.
Хотя идентификация функциональных изменений в регуляторных последовательностях остается серьезной задачей. эти две работы продемонстрировали мощь комбинированной биоинформатики и экспериментальных тестов. Однако большинство регуляторных элементов не являются высоко законсервированными среди видов и не состоят из кластеров одного и того же связывающего мотива. В самом деле, мало хорошо документированных случаев, связывающих некодирующие мутации с эволюционными признаками у людей и мух (7-9) с участием областей, которые были идентифицированы как функциональные на базе консервации последовательностей или кластирования мотивов.
Задачей на сегодня является разработка методов, которые распознают функциональные изменения внутри более значительной пропорции регуляторных элементов. Филогенетические выборки секвенированных геномов растут и становится возможным выявлятьбеспристратсно регуляторные изменения, базируясь на функциональных подходах на шкале генома (10), количественной генетике (11) и тестах по позитивному отбору (12).
|