Morpheus Unbound: Reimagining the Morphogen Gradient
Arthur D. Lander Cell. V.128, No.2, P. 245-256 | DOI 10.1016/j.celJ.2007.01.004
The theory that the spatial organization of cell fate is orchestrated by gradients of diffusing molecules was a major contribution to 20th century developmental biology. Although the existence of morphogens is no longer in doubt, studies on the formation arid function of their gradients have yielded far more puzzles than answers. On close inspection, every morphogen gradient seems to use a rich array of regulatory mechanisms, suggesting that the tasks carried out by such systems are far more extensive than previously thought.
Anna Kicheva, Periklis Pantazis, Tobias Bollenbach, Yannis Kalaidzidis, Thomas Bittig, Frank Julicher, Marcos Gonzalez-Gaitan Kinetics of morphogen gradient formation. Science 315, 521–525 (2007)
Секретируемые морфогены формируют концентрационные градиенты, которые предоставляют позиционную информацию во время развития; следовательно, концентрация и распределение морфогена д. тонко регулироваться. Но как контролируются дипазон и форма этих градиентов? Gonza'lez-Gaita'n с коллегами показали, что морфогены с разными диапазонами градиентов имеют разные механизмы и кинетики распространения через эпителий.
Авт. сравнивали два морфогена, Decapentaplegic () и Wingless (), оба они распространяются не направленно (non-directionally) в крыле Drosophila melanogaster, но с разными характеристиками распространения; Dpp формирует градиент более длинного диапазона, чем Wg. Чтобы исследовать, как контролируется диапазон градиента, они использовали fluorescence recovery after photobleaching (FRAP) функциональные green fluorescent protein (GFP)–Dpp или GFP–Wg слитые конструкции и измеряли 4 важные кинетические параметры, которые управляют распространением градиента: скорость продукции, коэффициент эффективной диффузии, неподвижная фракция и скорость деградации.
Величины 4-х кинетических параметров были отличными для Dpp и Wg. Напр., скорость деградации Wg была приблизительно в 5 раз выше, чем у Dpp, это может объяснить более короткий диапазон градиента Wg. Различные значения наблюдались тажке для неподвижной фракции этих молекул — 62% из Dpp молекул были неподвижными, тогда как Wg пул был практически полностью подвижным при 25°C.
Наконец, авт. проанализировали роль эндоцитоза на формирование градиента, используя температурочувствительный dynamin-mutant shibirets1, чтобы наблюдать острые эффекты блокирования эндоцитоза во FRAP эксперименте. Эндоцитоз оказался необходимым не только для деградации Dpp, но и для перемещения Dpp. Напротив, транспорт и деграция Wg не зависели от dynamin-обусловленного эндоцитоза.
Авт. пришли к выводу, что "different morphogen gradients can be generated by independent fine-tuning of the four important kinetic parameters ... and that different morphogens may use different mechanisms of transport and cellular machineries to achieve the formation of morphogen gradients." Дальнейший анализ необходим для измерения кинетики др. морфогенов, таких как Hedgehog, и для изучения, законсервированы ли механизмы транспорта морфогенов у разных видов.
FURTHER READING
Gonza'lez-Gaita'n, M. Signal dispersal and transduction through the endocytic pathway. Nature Rev. Mol. Cell Biol. 4, 213–224 (2003)
Проблема образования эмбрионального паттерна среди немногих давно стоящих проблем биологии является наиболее любопытной и источником существенных разочарований. Это проблема определения, как крупно-масштабная организация типов клеток в пространстве диктуется с помощью набора инструкций - генов - которые находятся в каждой клетке. Врачи наблюдают появление купно-масштабных признаков из мелко-масштабных элементов, компьютерщики наблюдают распределенную обработку информации: инженеры наблюдают мощные контрольные системы, а математики наблюдают соединенные частичные дифференциальные уравнения с интересным поведением.
За столетие исследований наиболее влиятельной концепцией в области формирования паттерна стала концепция морфогена. В старом смысле слова морфоген является субстанцией, которая продуцируется клетками и организует паттерн путем распространения по др. клеткам. Большинство современных биологов принимает более узкие определения в соответствии с частными теориями о том как работают морфогены. Мнение, превалирующее в экспериментальной литературе, начинается с Wolpert (1969), который предположил, что плавно снижающиеся градиенты, формируемые диффузией морфогенов от источника, снижаясь, наделяют позиционными значениями клетки, которые затем воспринимают разные судьбы в зависимости от назначенных им значений. Согласно этому мнению морфоген не является непосредственной инструктивной молекулой, а такой, которая дает качественно отличающиеся инструкции в зависимости от своей концентрации. Что же в конечном итоге предпоределяет паттерн, это градиенты морфогенов, превышающие пороговые значения, при которых гены включаются или выключаются.
Другая концепция морфогена исходит из работ Meinhardt and Gierer (Gierer and Meinhardt, 1972; Meinhardt and Gierer, 2000), которые расширили более ранние достижения Turing (1952). Эти работы показали, как два морфогена, которые влияют на синтез др. др., могут запускать спонтанное возникновение стабильных, дально-действующих паттернов активности морфогенов. При числовых моделированиях механизмы Meinhardt-Gierer продуцируют паттерны повторяющихся полос и точек, которые жутко напоминают некоторые из тех. что обнаруживаются в природе.
Воздействие этих теорий на биологию развития оказалось огромным, досих пор отношение к ним экспериментальных биологов нерушимо в лучшем случае. С годами неспособность идентифицировать какие-либо животные морфогены привело к широко распространившемуся сомнению, что такие субстанции существуют. С середины 1990s эта ситуация изменилась, благодаря результаты исследований на bicoid, внутриклеточном морфогене (Driever and Nusslein-Volhard, 1988), и Decaptentaplegic (Dpp), внеклеточном морфогене (Ferguson and Anderson, 1992; Nellen et al., 1996), оба из которых вносят вклад в развитие Drosophila. В последнюю декаду полипептидыиз семейств fibroblast growth factor (FGF), epithelial growth factor (EGF), Wnt, Hedgehog и transforming growth factor (TGF)-β, а также витамина А метаболит ретиноевая кислота, обнаружили поведение подтверждающее или сходное с морфогенами (напр., Green, 2002; Tabata and Takei, 2004; Schier and Talbot, 2005).
Несмотря на такой прогресс, отсутствует полный комфорт от концепции морфогена, сохраняющейся среди многих биологов. Некоторые предпочитают объяснять формирование паттерна на уровне сетей регуляторных генов, придавая минимальное значение движениям молекул, которые кодируются генами (напр., Davidson et al., 2002). Др. полагают, что градиенты морфогенов существуют, но возникает вопрос, адекватна ли диффузия и достаточно ли благонадежна, чтобы формировать паттерн (Kerszberg and Wolpert, 1998; Pfeiffer and Vincent, 1999).
В самом деле, нужна надежность - или как говорят инженеры прочность - в формировании паттерна, это становится навязчивой идеей среди экспериментаторов и теоретиков, ведущей многих к поискам свежих подходов к тому. как морфогены делают свою работу (Kerszberg, 1996; Pages and Kerridge, 2000; Pfeiffer et al., 2000; Belenkaya et al., 2004; Bollenbach et al., 2005). Хотя это и здоровое стремление, но существует некоторый непропорциональный акцент в отношении открытия новых молекулярных механизмов скорее, чем на полное эксплуатирование уже известных механизмов. Напр., не менее 4-х отдельных механизмов было предложено в качестве средств для (воспринимаемых) недостаточностей диффузии при транспортировании морфогенов от одного места к др. (Kerszberg and Wolpert, 1998; Ramirez-Weber and Kornberg, 1999; Entchev et al., 2000; Greco et al., 2001; Belenkaya et al., 2004; Kruse et al., 2004). Конечно же, импульс найти решения сложных проблем в новых механизмах вовсе не нов в науке. Вспомним убеждение Schrodinger's, что только новая квантовая физика сможет бъяснить постоянство генетического материала у живых существ (Schrodinger, 1944).
Мы можем спросить, действительно ли возможно пересмотреть существующие теории морфогенов и формирования паттерна, чтобы определить, до какой степени находки и проблемы современной экспериментальной биологии могут быть приспособлены и до какой новые механизмы следует искать. Эта цель, как я полагаю, может лучше соответствовать теории интегрированных морфогенов в широком контексте проблем пространственной динамики в биологии. Это могло бы также помочь дать более богатое, более полное описание того, что является требуемыми рабочими характеристиками систем формирования паттерна.
Space: A Final Frontier?
Жизнь является динамичной по многим временным рамкам. Молекулы соединяются и реагируют, клетки приходят и уходят, организмы рождаются и гибнут, виды развиваются. Молекулярные биологи построили солидную основу для понимания динамики процессов, лежащих в основе жизни, путем принятия концепций химии, таких как кинетика, термодинамика и сродство. Используя такие инструменты, в области, такой как метаболизм, экспрессию генов и внутриклеточную передачу сигналов, были исследованы сложные количественные пути.
Но жизнь является динамичной и отношении пространственной шкалы. Под этим я понимают, что материал жизни не является униформным в пространстве - "well-stirred," (хорошо перемешанным), используя химическое выражение - а устроен в сложных формах, от макромолекулярных ансамблей до мигрирующих стад. Вопросы, касающиеся пространственной динамики - как возникают структуры и паттерны и используются - важны для биологии, как и касающиеся временной динамики. Пространственная динамика д. покоиться на твердой количественной основе (инструменты более часто происходящие из физики, чем химии), но она требует значительно более нелегких усилий. Это частично из-за того, что пространство имеет три независимых измерения, а также из-за того, что объекты движутся взад и вперед (тогда как во времени вещи движутся только вперед) и потому, что интересующие пространственные вопросы часто затрагивают и пространство и время.
Математические, компьютерные и бухгалтерские скачки с препятствием, ассоциированные с описанием, анализом и моделированием пространственного феномена могут быть чрезвычайно затруднительны, особенно если пространственная организация не может быть приурочена к немногим хорошо очерченным компартментам. Проблемы подобного типа стремятся стать такими, в которых имеется скорее непрерывная, чем ступенчато-образная изменчивость признаков в пространстве, напр., такие, в которых имеют значение молекулярные градиенты. Формирование эмбрионального паттерна явно попадает в эту категорию, но также действуют и др. процессы, такие как контроль клеточного движения и формы с помощью хемоаттрактантов и взаимодействие внутриклеточной передачи сигналов с формой и структурой клетки. Прогресс в направлении количественного понимания ускорился в каждой из этих областей (напр., Iglesias and Levchenko, 2002; Wiley et al., 2003; Reas and Ballaro, 2004; Meyers et al.. 2006), частично благодаря улучшениям в скорости и мощи компьютеров. Изучение литературы указывает на то, что генеральные темы и стратегии, общие многим пространственно динамичным биологическим системам, начинают всплывать.
Walking the (Random) Walk
Красной нитью среди многих пространственно динамических проблем в биологии является то, что интересные поведения вытекают из случайно направленных движений молекул. В макроскопическом мире, типом движения, с которым мы обычно сталкиваемся, является то, что физики называют баллистикой: объекты остаются в покое до тех пор, пока на него не будет воздействовать сила и тогда они движутся в направлении, заданном силой, некоторое время пока в результате трения они не остановятся. Напротив, молекулы внутри и вокруг клеток находятся в постоянном движении с чрезвычайно высокими скоростями (результат тепловой энергии), но перемещаются только на очень незначительные расстояния, прежде чем столкнуться с др. объектами и случайно изменить направление (Berg, 1993). Т.к. траектория любой индивидуальной молекулы приближается к случайному блужданию, то ряд статистических правил может описать их коллективное поведение. Напр., если группа молекул, помещенная в одном месте, распространяется предсказуемым способом и заполняет остальное пространство. Такие молекулы перемещаются от высокой к низкой концентрации, создавая впечатление что различия в концентрации являются сортирующей силой ("двигающей силой"). Это позволяет предположить, имея в виду совокупность молекулярных движений, что и здесь может быть использована повседневная баллистическая интуиция. Это далеко от истины. Примем во внимание, что объекты, движущиеся баллистически, имеют скорость, тогда как объекты, распространяющиеся случайно, не имеют. Если требуется 10 мин для набора случайно движущихся молекул, чтобы переместиться на 10 um, то будет необходимо 40 мин, чтобы переместиться на 20 um и 90 мин, чтобы пройти 30 um (квадратическое взаимоотношение между временем и расстоянием является кардинальным свойством случайного блуждания). То что мы наблюдаем, как молекулы распространяются, не зависит от скорости, а от коэффициента диффузии (или diffusivity). Экстрагировать эти числа из экспериментальных данных трюкачество, особенно если молекулы не только перемещаются, но м также участвуют в связывании, деградации и химических модификациях.
В случае градиентов морфогена проблема связана не только с тем, что транспорт трудно измерить, но и с тем что применение баллистических представлений к экспериментальным данным легко ведет к недоразумениям. Замечательный пример приведен на Рис. 1А.
Figure 1. Consequences of Random, Diffusive Transport
(A) Concentration profiles for a morphogen that is continuously produced at constant rate for a fixed time. The solid and dashed curves display results for two different diffusion coefficients that differ by a factor of 5. For high-threshold morphogen responses (those induced when morphogen levels are above the red line), lower diffusivity shifts the response boundary farther from the morphogen source (red arrow). For low-threshold responses (blue line), the opposite occurs (blue arrow). Calculations assume that morphogen is produced by a stripe of cells (from -20 to 0 µm) within an effectively two-dimensional epithelium, so that results may be displayed as one-dimensional graphs. The expression used to calculate these profiles was obtained from Green's function for free diffusion by integrating over space and time (from 0 to 200 s). Diffusion coefficients for the two cases were 5 x 10~7 cm2 s-1 and 10-7cm2s-1. A very similar effect of diffusivity on gradient shape may also be observed for steady state gradients, depending on the conditions that produce the steady state (unpublished observations). (B and C) Simulations of ballistic and diffusive transport, and their responses to barriers. In both cases, 105 moving objects are released-10 per second for 10,000 s-from a single point into a field possessing an impenetrable barrier at one location (arrow). Object positions at the end of the time period are shown by individual dots (upper images), and via histograms of object density along the horizontal axes at the level of the barrier (lower images). In (B) ballistic motion is shown: the objects are tennis balls hit in random directions. Note the large number of balls that accumulate in front of the boundary. In (C) diffusive motion is shown: the objects are morphogen molecules undergoing random walks. Although the slope of the morphogen gradient flattens near the boundary, which may give a visual impression of slight accumulation, the histogram shows that there is no significant build up of molecules at that location.
Сплошная кривая является пространственным градиентом, который д. быть сформирован с помощью свободно диффундирующего морфогена с диффузионной способностью типичного белка, который постоянно продуцируется в течение 200 s в домене шириной 20-µm. Штриховая кривая показывает, что д. произойти в тех же самых условиях, если бы диффузионная способность морфогена снизилась в 5 раз. Если этот морфоген индуцирует определенный ген при пороге концентрации 8, то замедление его диффузии д. привести к уменьшению ширины домена индукции гена со 100 pm до 65 pm. Но если порог экспрессии гена равен 20, то домен индукции гена д. возрасти с 15 pm до 35 pm. Как же можно делая морфоген, движущимся более медленно, заставить его действовать дальше? Объяснение в том, что пониженная диффузионная способность позволяет морфогену накапливаться на более высоких уровнях вблизи своего источника. В недавней модели градиента sonic hedgehog (Shh) , который формирует паттерн вентральной части нервной трубки у эмбрионов кур (Saha and Schaffer, 2006), как раз возникает версия подобной ситуации, в которой подсчитано, что снижение диффузии Shh д. увеличивать диапазон действия Shh. Это является находкой чрезвычайной практической важности: согласно современному здравому смыслу heparan sulfate proteoglycans способствуют транспорту Drosophila hedgehog (Hh), , т.к. удаление их заметно снижает диапазон действия Hh (Bellaiche et al., 1998; Lin, 2004). Данные Saha and Schaffer (2006) говорят нам о том, что эти важные наблюдения д. указывать на протеогликаны как простейшее средство ингибирования транспорта Hh!
Др. не согласующимся с интуицией свойством случайного транспорта является его способ на препятствия. Рассматривая экспериментальные манипуляции, которые заставляют накапливаться морфогены в дискретных местах, таких как край клона мутантных клеток, обращенный лицом к источнику морфогене. Обычная интерпретация, что морфоген накапливается, т.к. он не может пройти сквозь клон (напр., Entchev et al., 2000). Это определенно ведет себя как баллистическое движение: если я стою на поле и ударяю по теннисному мячу во всех направлениях, то мячик будет приземляться с большим разбросом с плотностью, которая будет снижаться с расстоянием; если стенка помещена в одной точке поля, то мячики будут накапливаться по её фронту (Figure 1B). Но это не говорит о том, как ведут себя молекулы: если случайно движущиеся молекулы высвобождаются из точечного источника и наталкиваются на барьер, то они просто движутся от него прочь и вокруг препятствия (Figure 1C).
Способность обходить препятствие также объясняет, почему свободно диффундирующие молекулы могут пересекать любой случайный лабиринт за то же количество времени, которое им потребно для прохождения той же дистанции в свободном пространстве (Rusakov and Kullmann, 1998). Этот результат поможет нам принять то, что противоречит интуиции: что лабиринт из извилистых межклеточных пространств в большинстве тканей представляет собой незначительное препятствие для свободной диффузии морфогена. Ограниченное осознание этого факта возможно и объясняет, почкму любые морфогены часто изображаются перемещающимися исключительно внутри относительно беспрепятственных пространств на апикальных поверхностях эпителия (Pfeiffer and Vincent, 1999; Christian, 2000; Belenkaya et al., 2004; Lin, 2004). В действительности апикальный транспорт испытывает значительно больше трудностей, чем базо-латеральный, это обусловлено потенциалом массивного морфогена теряться в лежащей поверх среде.
Steady or Not, Here They Come
Странный мир случайно движущихся молекул кажется ещё более странным, если мы добавим эффекты продукции и деструкции. В хорошо-перемещающихся системах, когда способность деструкции субстанции превосходит скорость продукции, то динамическое равновесие может достигаться, при котором количество субстанции стремится в направлении постоянного значения. То же самое верно для пространственных динамических систем, за исключением тех, когда продукция и деструкция происходят в разных местах, формируются стабильные градиенты. Существует удобное правило большого пальца для таких градиентов. Напр., между дискретным источником и дискретным сливом д. формироваться линейный градиент. Напротив, когда диффузия от локального источника сбалансирована за счет деструкции, которая происходит с постоянной вероятностью повсюду, то устойчивый градиент имеет экспоненциальную форму. Длина, после которой такой градиент распадается до 1/e от своего наивысшего значения - число, иногда обозначаемое как шкала длины градиента - равная квадратному корню от соотношения диффузионной способности к скорости деградации, д.быть постоянной (Eldar et al., 2003; Gregor et al., 2005; Reeves et al., 2006). Длина любой интересующей области, подразделяемая с помощью масштаба длин (length scale) диффузии молекул в ней, является её Thiele modulus, термин недавно заимствованный из инженерии (Goentoro et al., 2006; Meyers et al., 2006).
Исторически большинство моделей градиентов морфогенов и формирования паттерна - взятых из "Wolpertian" или "Meinhardtian" перспектив - исходит из предположения, что паттерн управляется устойчивыми (steady-state) градиентами. В самом деле, кажется логичным, что информация о формировании паттерна должна быть до некоторой степени поддерживаемой стабильно, особенно из-за реакций нижестоящих клеток (напр., экспрессии генов), она д.быть относительно медленной по сравнению со временем, необходимым для диффузии молекул, чтобы перемещаться от одной клетки к др. В самом деле, для градиентов некоторых морфогенов, таких как градиенты Dpp и Wg в крыловых имагинальных дисках Drosophila, эксперименты показали, что время, в течение которого формируются градиенты, является более коротким, по сравнению с несколькими днями, в течение которых происходит формирование паттерна (Strigini and Cohen, 1999).
Но др. случаи менее ясны: у эмбрионов рыбок данио, только 4-5 ч проходит между поздней бластулой - когда возникают домены локальной экспрессии bone morphogenetic proteins (BMPs), Wnts, Nodal, FGFs и ретиноевой кислоты - и стадией поздней гаструлы, во время которой формируется огромое количество оркестрированных паттернов с помощью этих молекул (Schier and Talbot, 2005). У эмбрионов Drosophila, события происходят ещё быстрее: при 25°C, градиент bicoid формируется и начинает действовать по формированию передне-заднего паттерна менее чем через 2 ч (Gregor et al., 2005), a градиент морфогена BMP morphogen на дорсальной срединной линии формируется и специфицирует дорсо-вентральный паттерн в течение 1 ч (Dorfman and Shilo, 2001).
Могут ли такие градиенты достигать устойчивого состояния достаточно быстро? Для большинства простых сценариев, доминантный фактор, предопределяющий скорость приближения к устойчивому состоянию является средней продолжительностью жизни молекулы морфогена (т.е., постоянно обратной по отношению к скорости её деградации [Gregor et al., 2005; Lander et al., 2005]). Для bicoid, внутриклеточного белка, продолжительность жизни порядка минут наиболее правдоподобна. Конечно, как аргументируют Gregor et al. (2005), т.к. скорость деградации также влияет на установившееся состояние масштаба длин (see above), то необходимо достижение устойчивого состояния в течение 1-2 ч что д. вынуждать градиенты bicoid достигать максимального размера в 1-2 mm (интерсно бы посмотреть насколько он велик по сравнению с эмбрионами насекомых).
Для секретируемых полипептидных морфогенов, деградация обычно осуществляется посредством последовательных ступеней связывания, обеспечиваемого рецепторами эндоцитоза и лизосомного протеолиза (Scholpp and Brand, 2004; Marois et al., 2006). Неясно, достаточно ли быстры такие события для формирования устойчивого градиента BMP у ранних эмбрионов мух в течение менее 1 ч. В самом деле, идея что этот градиент не формирует паттерна до тех пор, пока не будет достигнуто, подтверждена экспериментально, т.к. обнаружены комбинированные фенотипы определенных мутаций, которые трудно объяснить др. способом (Mizutani et al., 2005). Др. случаи, при которых анализ подтверждает, что устойчивые состояния не достигаются с помощью градиентов морфогена, включают ранее упомянутую модель формирования паттерна нервной трубки с помощью Shh (Saha and Schaffer, 2006) и некоторые модели сети сегментации у эмбрионов Drosophila (Gursky et al., 2004).
Почему важно знать, происходит ли формирование паттерна с помощью градиентов морфогена в меняющихся или устойчивых состояниях? С одной стороны, лежащая в основе математика говорит нам, что реакции градиентов на внешние воздействия могут быть очень различными при двух режимах. Попытаемся предсказать эффекты "неспецифических" сайтов связывания: если диффундирующий морфоген соединяется обратимо с неподвижными сайтами в своей среде, тогда в пространстве, в котором он это осуществляет, он распространяется более медленно. В каждый момент времени во время достижения устойчивого состояния, наблюдаемый градиент д. быть более узким (из-за снижения способности к диффузии). Но в устойчивом состоянии профиль градиента д. оставаться тем же самым, как если бы не было неспецифических сайтов связывания (достигнутое локальное равновесие между захватом и высвобождением достигается повсюду, оно не оказывает влияния на течение морфогена [Eldar et al., 2003]). С др. стороны, изменения нетто коэффициента деградации морфогена всегда д. влиять на форму градиента в устойчивом состоянии (масштаб длины обратно пропорционален квадратному корню из константы скорости деградации), но он может оказывать лишь незначительные эффекты во время достижения устойчивого состояния, особенно в ранний период и вблизи источника морфогена.
Соответственно, должен ли исследователь интерпретировать экспериментально индуцированные изменения в форме градиента морфогена, как доказательство более или менее неспецифического связывания, как большую или меньшую степень деградации, более быструю или медленную диффузию или любую изменчивость каждой из альтернатив, серьезно зависит от того, сделаны ли наблюдения в устойчивом состоянии, и в некоторых случаях от того, где в градиенте оно отмечено. Простые экспериментальные манипуляции, обычно затрагивают более одного важного аспекта формирования градиента во времени. Напр., добавление растворимого, связывающего морфоген белка в систему устойчивого градиента морфогена д. обнаруживать тенденцию к глобальному уменьшению передачи сигналов морфогена (путем уменьшения овладения рецепторов морфогена), но может также существенно увеличивать шкалу длины градиента (путем снижения скорости обусловленной рецепторами деструкции морфогена). Суммарное воздействие может быть суммарным снижением передачи сигналов морфогена в некоторых местах, связанным с суммарным увеличением в других (Mizutani et al., 2005).
Complexity and Performance
Тот факт, что некоторые градиенты морфогена могут делать свою работу только в условиях неустойчивых состояний как раз является одним из указаний на то, что формирование паттерна с помощью морфогена более сложное, чем мы привыкли думать. В самом деле, в последнюю декаду, открыт удивительный набор регуляторных механизмов для широкого диапазона систем градиентов морфогена.
Некоторые из них, по-видимому, непосредственно связаны с определенными свойствами случайного транспорта. Напр., морфогены из семейств hedgehog и Wnt часто модифицируются липидами, это указывает на путь контроля из диффузионной способности (Eaton, 2006). Секретируемые ингибиторы широко используются в градиентах BMP, activin, nodal, Wnt и EGF (Kawano and Kypta, 2003; De Robertis and Kuroda, 2004; Klein et al., 2004; Yamamoto et al.,c2004). Неспецифические (или более правильно, не рецепторные) сайты связывания для BMPs, Wnts, hedgehogs и FGFs обнаруживаются в изобилии в форме heparan sulfate proteoglycans клеточной поверхности, а для ретиноевой кислоты в форме клеточных, связывающих retinoid белков (Ross, 1993; Lin, 2004).
Доказательства для сложной регуляции не прекращаются: почти каждый морфоген взаимодействует с одним или более типом ко-рецепторов, приблизительно обозначаемых как белок, связывающий морфоген клеточной поверхности, который усиливает образование комплексов морфоген-рецептор и/или усиливает передачу ими сигналов (Kirkbride et al., 2005). В некоторых системах градиентов пары морфогенов действуют в содружестве, иногда взаимно усиливая др. др. (e.g., Nguyen et al., 1998). Для некоторых морфогенов, ограниченных транспортными путями, градиенты могут существовать вдоль клеточных структур, таких как cytonemes (Ramirez-Weber and Kornberg, 1999), или за счет внутриклеточных челноков (Greco et al., 2001).
На вершине этого, выявляемый экспериментальной работой, богатый набор механизмов обратной связи, управляемых с помощью передач сигналов морфогенов. Примеры, в которых передача сигналов морфогена или усиливает или ослабляет дальнейшую передачу сигналов морфогена, продукцию морфогена, деструкцию морфогена, синтез рецепторов морфогена или синтез ко-рецепторов морфогена обнаруживаются в исследованиях по формированию паттернов с помощью TGF-βs, hedgehogs, Wnts, EGFs, FGFs и ретиноидов (e.g., Cadigan, 2002; Green, 2002; Fujise et al., 2003; Dobbs-McAuliffe et al., 2004; Lai et al., 2004; Ben-Haim et al., 2006). В самом деле, обнаруживаются очень редко системы морфогенов, которые лишены обратной связи.
Когда впервые появилась классическая модель градиентов морфогена Wolpert и Meinhardt, то недостаток механистической информации заставил сформулировать минимальные описания систем формирования паттерна. Учитывая насколько хорошо эти модели работают в теории, кажется неожиданным, что существуют столь множественные дополнительные сложности in vivo. В биологии находка неожиданного усложнения обычно означает, что существуют ограничения того, как система оперирует, о которых мы не подозревали. Хотя некоторые ограничения могут быть наложены на градиенты морфогенов за счет физических и химических ограничений (Lander et al., 2002), большинство несомненно накладывается за счет эволюции. Благодаря естественному отбору градиенты морфогенов, наблюдаемые в мире сегодня, являются теми, которые выполняют работу особенно хорошо. Это особенно верно для систем градиентов морфогенов, которые были законсервированы на больших эволюционных дистанциях.
Если системы морфогенетических градиентов усложняются, чтобы лучше осуществлять свою функцию, то ключом к открытию их сложности, как известно, является то, как в действительности выполняется работа. На первый взгляд эта система тривиальна: морфогены создают паттерн. И всё же в противоположность этим поддающиеся определению единицам - назовем их требуемые рабочие характеристики - мы наталкиваемся на разнообразие нетривиальных индивидуальных целей: встречаем временные спецификации, делающие четкими границы, предназначения множественных судеб, тотчас продуцирующие периодические паттерны, связь паттернов др. с др. и т.д. Кроме того, исполнение таких целей реально перед лицом мириад средовых и генетических пертурбаций д. непременно быть чем-то таким, по чему происходит естественный отбор. Т.о., необходимо включать выносливость-резистентность к пертурбациям - в качестве необходимых рабочих характеристик. Однако, из-за средовых изменений, которые могут влиять на скорость, с которой происходит общее развитие, и на размеры структур с формируемыми паттернами, градиенты морфогенов д. временами приспосабливаться скорее к, чем противостоять, изменениям (и имеется набор экспериментальных доказательств, что они действуют [напр., Teleman and Cohen, 2000]). Адаптивность, как необходимая рабочая характеристика, возникает в др. контексте: т.к. изучаемые организмы являются производными родоначальников очень разных форм, размеров и онтогенетического временного графика, и только такие градиенты морфогенов способны приспособиться к соотв. наблюдаемым отличиям; такие необходимые рабочие характеристики иногда обозначают как способность к развитию (evolvability) (Meir et al., 2002).
Проблема, изложенная с помощью объективных рабочих характеристик, как раз не является той, которых множество, а той стратегией, которая поддерживает один частый компромисс другим. Это находится в соответствии с таким выбором оптимальных соотношений, которые необходимы для сложной регуляции, находящейся в центре внимания. Этот вопрос постоянно возникает перед инженерами: построить автомобиль, который легко ускоряется, поворачивает и останавливается. Чтобы сделать подобный, он д. обладать множеством др. необходимых рабочих характеристик - управляемостью, надежностью, видом, ценой, долговечностью и т. д. Это в свою очередь служит поводом для добавления свойств, подобных рулевому управлению с гидроусилителем, воздушным подушкам, комбинированному индукторному двигателю, бортовому компьютеру и т. д. Сегодня только часть из всей этой кухни в среднем автомобиле действительно непосредственно участвует в ускорении, остановке и поворотах. Большая часть оставшегося представлена мириадами контрольных систем, важность которых может быть раскрыта только при специальных условиях (напр., воздушные подушки при столкновении).
Нет причин ожидать, что организм отличен от автомобиля в этом отношении, конитрольные системы, по крайней мере, настоль же многочисленны, что механизмы, выполняющие основные функции. Вот почему Doyle активно аргументирует (Csete and Doyle, 2002; Stelling et al., 2004), инженерными концепциями и теориями, особенно теми, что вытекают из изучения контрольных систем, пытаясь ввести их в биологию. К сожалению, для тех из нас, работающих с морфогенетическими градиентами, основная масса этой теории была разработана с помощью временных, а не пространственных, динамических систем. Позитивной стороной этого умозрения является то, что изучение систем морфогенетических градиентов может рассматриваться как передний край ("cutting edge") как для биологов, так и инженеров!
Figure 2. Strategie for Making Sharp Borders (MD) Each panel shows calculated shapes of signaling gradients pro-duced at three rates of morphogen production, over a field of 20 rows of cells, using different sharpening strategies (units of signaling are arbitrary). The shaded box gives the distance from the tenth row to the location where the middle curve falls by half. The width of the box thus reflects the steepness of the curves, whereas the distance between curves reflects robustness to variations in morphogen synthesis rate. The shaded box of panel (A) is extended as a Iight stripe into panels (B)-(D) to facilitate comparison. In (A), no sharpening strategy is shown. The morphogen gradient is a declining exponential, and intracellular signaling is proportional to the amount of extracellular morphogen. In (B), the morphogen gradtant is the same as in panel (A), but there is positive feedback in intracelular siynanig (a key signaling molecule inhibits its own destruction), In (C), morphogen synthesis rates are high enough that receptors are saturated near the start of the gradient. In (D), morphogen stgnafng upregulates expression of morphogen receptors. In (A), (C), and (D), the black, blue, and red curves represent successive 2-fold increases in morphogen synthesis rate, In (B), these curves represent only 8% increases. Notice that in (B), steep declines in signaling over one cell row are easty obtained, but tolerance for variation in morphogen synthesis rate is drarnabcaiy reduced. In (C) and (D), robustness to morphogen synthesis rate is improved, but other performance objectives are compromised (not shown).
Trouble at the Border
Имеется множество способов, чтобы начать углубляться в вопрос объективных рабочих характеристик и выбора оптимальных соотношений в системах морфогенетических градиентов. Сначала рассмотрим наиболее частую цель формирования онтогенетического паттерна: организацию клеток в дискретные домены экспрессии генов с рассыпчатыми границами (rev. Irvine and Rauskolb, 2001). Чтобы понять, что это влечет за собой, представим картину, что вы задумались над планом создать из собранного наспех систему морфогенетического градиента, которая бы достигала этой цели.
Чтобы начать вы можете выбрать классический Wolpert-типа градиент, в котором морфоген продуцируется в одном месте и просто транспортируется прочь с помощью диффузии. Предполагая, что морфоген является секретируемым полипептидом, соединение с высоким сродством с рецепторами на клеточной поверхности может обеспечить передачу сигнала и заботиться об удалении морфогена. До тех пор, пока морфоген продуцируется достаточно медленно, чтобы рецепторы не оказались насыщенными, будет возникать устойчивый экспоненциальный градиент и морфогена и оккупированных им рецепторов. До тех пор, пока размер поля, которое д. подвергнуться формированию паттерна, не является достаточно большим, достижение устойчивого состояния оккупации рецепторов д. происходить в то же самое время, которое используется рецепторами, связавшими молекулы морфогена, чтобы быть разрушенными.
Предположим такой градиент формирует паттерн поля шириной в 20 клеток и вы желаете предопределить положение четкой границы генной экспрессии между 10-м и 11-м рядом клеток. Если градиент имеет типичный Thiele модуль из ~3 (Goentoro et al., 2006) (т.е. скажем градиент снижается на e-3, или до 5%, от инициального значения к 20-й клетке), то различия в оккупации рецепторов морфогеном между двумя соседними рядами клеток д.б. более 16% (Figure 2A). Если нижестоящая передача сигналов пропорциональна оккупированности рецепторов, то вы будете иметь проблему, т.к. экспериментальные исследования показали, что 2-кратные или большие изменения в передаче сигналов необходимы, чтобы продуцировать резкие различия в экспрессии генов (Shimizu and Gurdon, 1999). Поэтому важно, чтобы трансформация мелководного морфогенетического градиента в острую границу генной экспрессии не была упущена биологами развития (e.g., Small et al., 1992; Dyson and Qurdon, 1998; Jaeger et al., 2004; Ashe and Briscoe, 2006). Т.к. это случилось, то имеется множество способов обойтись с этим.
Напр., вы можете заменить линейные взаимоотношения между оккупацией рецепторов и передачей сигналов со степенью 1. Одним из механизмов сделать это является введение позитивной петли обратной связи в путь передачи сигналов - , напр., позволяя морфогену ингибировать деградацию компонента этого сигнального пути. При правильных условиях клетки в ряду 10 легко смогут воспринимать в 5 раз сигнала по сравнению с клетками в ряду 11 (Figure 2B). Или вы можете сделать передачу сигналов высоко кооперативной - напр., с помощью привлечения димеризации сигнальных молекул или кооперативного действия транскрипционных факторов на их промоторы. Или, вы можете комбинировать как негативную, так и позитивную петли обратной связи, чтобы генерировать феномен, известный как бистабильность, так чтобы передача сигнала морфогена резко прыгала с одного потенциально устойчивого состояния к другому в специфической точке градиента (Von Dassow and Odell, 2002; Ingolia, 2004; Lai et al., 2004). Или, вместо только использования передачи сигналов морфогена, чтобы управлять активацией генов, вы можете использовать его использовать для превращения репрессора в активатор, получая два эффекта за одну цену. Вы можете также воспользоваться преимуществами феномена, известного как zero-order ultrasensitivity (Goldbeter and Koshland, 1981), при котором два антагонистических ферментативных процесса оперируют в условиях высокого насыщения энзимами, приводя к тому, что исход системы становится чрезвычайно чувствительным к соотношению уровней двух энзимов. Вы можете даже эксплуатировать феномен, известный как стохастическое фокусирование (Paulsson et al., 2000), в котором чувствительное поведение возникает в результате статистических эффектов, которые становятся достоверными, когда количество сигнальных молекул на клетку мало.
Приведенные выше механизмы несмотря на их различия, дают преимущества только временной, но не пространственной, динамике. Принимая более экспансивную точку зрения вы позволите себе открыть программу из дополнительных стратегий. Некоторые являются пространственными аналогами временного феномена. Напр., вы можете иметь сначала размытые границы, которые сужаются сами по себе, благодаря способности клеток перестраивать свои позиции, или меняя их генную экспрессию, исходя из сравнений между их реакциями и реакциями их соседей (напр., Rogulja and Irvine, 2005). Если такой процесс перемещает клетки со сходными реакциями ближе др. к др., то эффект будет по существу одним из кооперативных; т.е., стабильные позиции клеток зависят от того, является ли их сложность сходной, подобной отвечающим клеткам.
Феномен напоминает ультрачувствительность нулевого порядка (zero-order ultrasensitivity), но применим к пространству скорее, чем ко времени, и может быть достигнут за счет регулировки скорости продукции морфогена, достаточно высокой, чтобы насытить рецепторы на части поля, в котором формируется паттерн. Такая стратегия может давать ступенчатое снижение передач сигналов морфогена в любом случайном местоположении (Figure 2C). В отличие от любой др. стратегии, рассмотренной выше, эта (также как и те, что будут рассмотрены ниже) работают путем изменения формы собственно градиента морфогена, а не только характеристик клеточной реакции.
Если это происходит, то осуществляемые эффекты обратной связи в пространстве, как вы скорее всего обнаружите, быстро осложняются. Рассмотрим морфоген, который позитивно регулирует синтез своего собственного рецептора (Figure 2D). Т.к. передача сигналов усиливается, когда имеется множество рецепторов, то это позитивная обратная связь. Но если рецепторы выполняют задачу деградации, тогда большое количество рецепторов означает, что морфоген сможет распространиться на меньшее расстояние; поэтому позитивная регуляция рецепторов будет также негативной обратной связью. Можно ожидать, что эти противоположные эффекты будут аннулироваться, но этого не происходит, т.к. каждый имеет разный пространственный диапазон. В ответ на увеличение количества рецепторов усиливается передача сигналов только в клетках, которые обладают добавочными рецепторами, но происходит депрессия передачи сигналов на расстоянии (меньшее количество морфогена делает это в др. клетках). Эта комбинация коротко-действующей активации и дально-действующего ингибирования может ударить по нервам кого-нибудь, хорошо знакомого с классической моделью морфогена Meinhardt и Gierer, которая использует те же самые производящие принципы для формирования четких полос и пятен. Традиционно, организация таких моделей требует пар антагонистических морфогенов с разной диффузионной способностью, но обсуждаемая выше предполагает, что сходное пространственно динамическое поведение может возникать за счет др. механизмов, даже с вовлечением одиночного морфогена.
Финальный пространственный механизм для продукции подобных границам профилей передачи сигналов морфогена, который заслуживает упоминания из-за своей явной искусности. В этой стратегии, которая вышла на свет благодаря результатам экспериментов с градиентом BMP, который формирует паттерн дорсо-вентральной оси у ранних эмбрионов Drosophila, диффундирующий BMP ингибитор Sog используется, чтобы переносить морфоген поверх своего собственного концентрационного градиента, где Sog расщепляется и высвобождается морфоген (Holley et al., 1996; Shimmi and O'Connor, 2003). Формально, это пример облегчения транспорта, где потенциальная энергия одного молекулярного концентрационного градиента используется для осуществления энергетически неблагоприятного транспорта другого.
Sharp, Prompt, or Reliable?
Из различных теоретических механизмов, описанных выше, каждый наблюдается в одной системе морфогенетического градиента или другой. В случае градиентов Hedgehog мы находим позитивную обратную связь, превращение ингибирования в активацию транскрипции, бистабильность и позитивный контроль синтеза рецепторов (Saha and Schaffer, 2006). При передаче сигналов ретиноевой кислоты мы также видим позитивную обратную связь и превращение ингибирования в активацию транскрипции, а также кооперативность (Kerszberg, 1996). Dpp превращает транскрипционное ингибирование в активацию косвенно, посредством подавления репрессора brinker, который может в некоторых случаях также управлять позитивной обратной связью (Jazwinska et al., 1999). Bicoid один из множества транскрипционных факторов, который действует кооперативно на свои транскрипционные мишени (Burz and Hanes, 2001). При градиенте EGF, который формирует четкие паттерны вентральной эктодермы у эмбрионов Drosophila, недавние доказательства подчеркивают использование zero-order ultrasensitivity (Melen et al., 2005). Доказательства для пространственной кооперативности за счет клеточной перестройки существуют в нескольких системах (Irvine and Rauskolb, 2001). А механизм облегченного транспорта BMP, упомянутый выше, по-видимому, используется только у эмбрионов мух и только при формировании крыловых жилок (O'Connor et al., 2006), и для формирования дорсо-вентрального паттерна у эмбрионов позвоночных (De Robertis and Kuroda, 2004).
Почему используется такое множество механизмов для выполнения одной и той же работы? Напр., некоторые стратегии, такие как те, что базируются на позитивной обратной связи и zero-order sensitivity, обнаруживают тенденцию к увеличению времени, необходимого для достижения устойчивого состояния, что может быть проблематичным, если важно быстрое формирование паттерна. Др. стратегии, такие как облегчение транспорта BMP, могут быть очень быстрыми, но имеются, по-видимому, ограничения в точности границ, которые они формируют (Umulis et al., 2006).
Необходимая рабочая характеристика, которая может быть легко поставлена под угрозу при всех стратегиях формирования границ, это эксплуатационная надежность (robustness). Это из-за того, по существу, что формирование четкой границы, по-видимому, связано с увеличением чувствительности: чувствительности передачи сигналов от уровня морфогена или чувствительности уровня морфогена от позиции. Напротив, надежность д. сопровождаться снижением чувствительности: чувствительности к вариациям в архитектуре системы, средовым пертурбациям или сигнальным шумам. Хитрость, как увидим, заключается в создании высокой чувствительности, когда подходящая и низкая чувствительность во всем остальном. Это не маленькая уловка. Независимо от выбора механизма формирования границы, обычно можно найти некоторый аспект надежности, который тяжело нарушен. Позитивная обратная связь, напр., легко делает хрупкость (противоположность надежности) параметром изменчивости (in Figure 2B, не более 8% изменений в синтезе морфогена заставляет границу в ряду 11 сдвигаться за пределы 19). Комбинация негативной и позитивной обратной связи, которая вызывает бистабильное поведение может уменьшать такую хрупкость, но не полностью (Saha and Schaffer, 2006). Стратегии, подобные zero-order ultrasensitivity, могут обнаруживать внушительную надежность к определенным вариациям (напр., к уровню насыщенной киназы или её фосфорилированного субстрата [Melen et al., 2005]), но только из-за того, что хрупкость заключена где-то в др. месте. Для многих из пространственных стратегий, рассмотренных выше, исчерпывающий анализ чувствительности не проведен, и всё же всё ещё легко обнаружить примеры существенной хрупкости.
К счастью, биология не требует надежности во всём (и это хорошо, т.к. инженерные аргументы говорят о том. что такая цель недостижима). Естественный отбор, как можно ожидать, делает биологические системы надежными к пертурбациям, с которыми они встречаются в природе. Эволюционное давление, связанное с любой из данных хрупкостей, д. быть сбалансировано между частотой появления пертурбации и нарушением приспособленности, ассоциированным с ошибкой, чтобы её компенсировать. Согласно этому аргументу, мы д. ожидать, что системы морфогенетических градиентов, которые используют разные морфогены, затрагивают события с разными временными шкалами, территории формирования паттерна разных размеров или оперируют у видов, которые населяют разные окружающие среды, должны отбираться на надежность разными путями.
Рассмотрим, напр., retinoic acid (RA), морфоген возникающий в результате ферментативной модификации витамина А. Т.к. витамин А получается с пищей, а диеты могут быть очень отличными, то можно ожидать, что величины синтеза RA являются довольно ненадежными. Соответственно, мы можем ожидать существование строго отбора для механизмов, которые делают градиенты RA надежными на уровне синтеза RA. Один такой механизм, реализация контроля негативной петли обратной связи, д. существовать в клетках, которые заставляют RA подавлять экспрессию энзимов биосинтеза RA в ответ на передачу сигналов RA. Альтернативная стратегия, которая использует обратную связь, но эксплуатирует также пространственную динамику, возможна для RA чтобы индуцировать, в RA-чувствительных клетках энзимы, которые деградируют RA (такая само-усиливающаяся деградация способствует надежности градиентов морфогена в отношении вариаций в продукции морфогена, уже была Интересно, что доказательства на базе систем формирования паттерна с помощью RA указывают на то, что обе стратегии обратных связей исполняются повседневно (Dobbs-McAuliffe et al., 2004).
Fueling New Thoughts
Мы можем предположить, что в каждой системе морфогенетических градиентов наблюдаемые нами динамичные и регуляторные механизмы непосредственно связаны с балансом необходимых рабочих характеристик, сриди них надежность в отношении некоторых, но не всех пертурбаций. Сходным образом все гипотезы, которые исходят из эволюционной теории, иногда могут оказываться ложными - некоторые сложные состояния дел в биологии могут быть историческими несчастными случаями. До сих пор такие гипотезы могли заставлять нас думать в направлении, в котором мы никогда бы не пошли.
Рассмотрим, напр., формирование паттерна нервной трубки с помощью Shh. Модель, представленная Saha and Schaffer (2006) генерирует четкие границы передачи сигнала примерно в 70 µm от источника морфогена в любую сторону, из которых специфицируются клеточные судьбы V3 интернейронов и мотонейронов. Формирование границы связано как с врожденной бистабильностью пути передачи сигналов Hedgehog, плюс с пространственным эффектом коротко-действующей активации, так и с дально-действующим ингибированием, ассоциированным с позитивной регуляцией рецепторов. Благодаря позитивной обратной связи система в течение длительного времени достигает устойчивого состояния, дольше фактически, чем временное окно, в течение которого специфицируются судьбы клеток. К счастью, бистабильность достигается ещё до достижения стабильного состояния и в результате фиксированная граница клеточных судеб устанавливается задолго до сигнальных различий, которые их стабилизируют (Saha and Schaffer, 2006).
Хотя надежность этого механизма ещё не изучена до глубины, более ранние работы той же самой группы подтвердили, что локализация границы скорее всего чувствительна ко многим параметрам (Lai et al., 2004). Поэтому можно предположить, что система обладает дополнительными петлями контроля, которые купируют нежелательный момент границы, чтобы откорректировать изменение по какому-нибудь параметру, затрагивающему границу. Такие петли не подтверждены в литературе, но имеются указания, если их поискать. Напр., моделирование и эксперименты указывают на то, что изменения в уровнях дополнительных молекул, таких как heparan sulfate proteoglycans и hedgehog-interacting-protein (Hip), могут использоваться для сдвига позиции границы в любом направлении и оба типа молекул, как известно, являются транскрипционными мишенями для Hh/Shh (Chuang and McMahon, 1999; Fujise et al., 2003; Saha and Schaffer, 2006).
Второй пример системы морфогенетического градиента, из которого могут возникнуть новые гипотезы на базе исследований необходимых рабочих характеристик, является градиент BMP, который формирует паттерн дорсо-вентральной оси у дрозофилы. Как упоминалось раньше, этот градиент использует облегченный транспорт, чтобы создать острый пик передачи сигналов BMP в дорсальной срединной линии эмбриона. Моделирование подтвердило, что надежность ширины этого пика является требуемой рабочей характеристикой, которая оправдывает эту сложную транспортную схему (Eldar et al., 2002), но наблюдения in vivo не подтверждают это мнение. В частности, ширина сигнального пика действительно очень хрупка по отношению к дозе Sog (Mizutani et al., 2005). Более того, экспериментальные условия, которые существенно меняют ширину пика, в общем-то дают нормально выглядящих мух, подтверждая, что отбор на высокую надежность пика не столь строг.
Чио же тогда является требуемой рабочей характеристикой, облегчающей транспорт? Возможная информация получена в недавнем наблюдении, что процесс позитивной петли обратной связи инициируется поздно в процессе формирования паттерна с помощью ген-регуляторных эффектов BMPs, это заметно заостряет сигнальный пик на срединной линии (Wang and Ferguson, 2005). Механизм, лежащий в основе этого процесса, который связан с бистабильностью градиентов Hh, неизвестен, хотя возможные модели предложены (Umulis et al., 2006). Мы можем дать рациональное объяснение снижению необходимости в надежности облегченного транспорта на основании того, что последующее, управляемое обратной связью событие независимо диктует паттерн. Это, однако, только голословное утверждение, почему облегченный транспорт необходим в первую очередь. В самом деле, формирование крыловых жилок у куколок - др. система формирования паттерна, в которой облегченный транспорт BMP, по-видимому, купирован с активацией петли позитивной обратной связи с формированием узких полос генной экспрессии (O'Connor et al., 2006) - мутации, которые преимущественно устраняют облегченный транспорт (Yu et al., 1996; Vilmos et al., 2005) оказывают незначительный эффект на ширину жилок (подразумевая, что тонкие полоски активности BMP могут создаваться без облегченного транспорта).
Значение того, что действительную рабочую характеристику облегченного транспорта BMP ещё предстоит установить, заставляет нас думать в новых направлениях. Напр., в очень быстрых условиях формирования паттерна у эмбрионов мух, вполне правдоподобно, что бистабильность, управляемая позитивной петлей обратной связи, слишком медленна, чтобы совершать работу. Вообще-то эмбрионы разрешают эту проблему прежде всего за счет использования очень быстрого облегченного транспорта, чтобы создать грубое приближение финального паттерна, после чего система позитивной обратной связи может начать свою работу с инициальными условиями, которые уже приблизили к желанной цели. Достижение как скорости, так и аккуратности за счет последовательного использования низкосортного, а затем тонкого контроля является стратегией, обычно используемой в инженерии, так что имеется резон ожидать встретить это и в мире клеток.
И всё же даже это объяснение скорее всего неполное. Как обсуждалось ранее, облеченное перемещение BMP управляется с помощью повторяющихся соединений и расщеплений Sog. У Drosophila, Sog расщепляется значительно быстрее, если соединен с BMPs (Shimmi and O'Connor, 2003), это вносит позитивную обратную связь в деструкцию Sog (расщепление Sog высвобождает BMPs, которые затем соединяются с Sog, заставляя его расщепляться). Такая петля обратной связи не нужна для облегченного транспорта per se, но она, как полагают, имеет интересный эффект на создание на дорсальной срединной линии пика BMP, который выглядит очень крутым (Mizutani et al., 2005), это согласуется с экспериментальными наблюдениями (Ross et al., 2001). Фактически система, по-видимому, обладает встроенным выключателем временной задержки. Может ли это быть некой рабочей характеристикой? Вообще-то имеется необходимость координации формирования дорсо-вентрального паттерна с др. событиями у ранних эмбрионов, такими как формирование передне-заднего паттерна или ядерные деления. Шаг из этого к др. быстро-двигающемуся процессу определенно чувствителен к средовым условиям (напр., температуре), так что вообще-то время задержки используется, чтобы приспособить переход формирования дорсо-вентрального паттерна к конкуренции. Если это так, то следует проследить механизмы, с помощью которых события формирования передне-заднего паттерна или цикла ядерных делений, снабжают сырьем только те параметры, которые прогнозируют контроль продолжительности временной задержки (такие как отношение скоростей продукции Dpp и Sog [Mizutani et al., 2005]).
Toward a Systems Biology of Pattern Formation
На предшествующих страницах я использовал отобранные примеры для обсуждения многих регуляторных механизмов, обнаруженных в системах морфогенетических градиентов, взаимоотношений этих механизмов с феноменом пространственной динамики и пригодности концепции необходимых рабочих характеристик для объяснения, почему системы морфогенетических градиентов конструируются такими, какие они есть. Я не стремился представить исчерпывающее или даже сбалансированное мнение обо всем, что известно о морфогенетических градиентах. Вместо этого я попытался использовать примеры литературы, чтобы проиллюстрировать, как некоторые из сложностей систем морфогенетических градиентов могут быть вычленены и до некоторой степени поняты.
Многие исходят из одной и той же точки зрения, используя разные системы морфогенетических градиентов. Напр., объективные характеристики точного времени и координации д. быть внесены в обсуждение из сомитных "clock" позвоночных (Dubrulle and Pourquie, 2004). Бистабильность необходимо обсуждать с учетом образования дорсальных придатков в яйцах насекомых (Shvartsman et al., 2002) или сети сегментной полярности у насекомых (Von Dassow and Odell, 2002; Ingolia, 2004). Дополнительные объективные характеристики д. быть внесены, такие как сильная амплификация едва-обнаружимых различий (ключевая ступень формирования лево-правостороннего паттерна [Nonaka et al., 2005]), супрессия пространственных шумов (Ashe and Briscoe, 2006) или координация паттернов с ростом (Lawrence, 2001).
Некоторые читатели могут быть смущены появлением терминов с телеологическими обертонами, таких как работа, цель и необходимые рабочие характеристики. В современной биологии (особенно молекулярной биологии) предположения, что объекты "предназначены" совершать работу или имеют "причины", имеют тенденцию будить подозрения в антиэволюционных взглядах . Фактически, напротив, это эволюция наделяет биологию телеологическим языком (Ayala, 199; Lander, 2004). Временами сказать о совершении определенной работы более естественно и менее окольно, чем сказать её структура поддерживается с помощью естественного отбора благодаря её способности вносить вклад в приспособленность вида определенным способом.
Др. взгляд: в последние годы биологи всё чаще стремятся помещать проблемы на системный уровень...

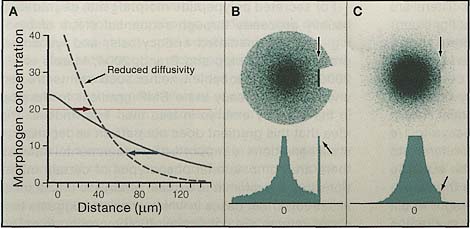 Figure 1. Consequences of Random, Diffusive Transport
(A) Concentration profiles for a morphogen that is continuously produced at constant rate for a fixed time. The solid and dashed curves display results for two different diffusion coefficients that differ by a factor of 5. For high-threshold morphogen responses (those induced when morphogen levels are above the red line), lower diffusivity shifts the response boundary farther from the morphogen source (red arrow). For low-threshold responses (blue line), the opposite occurs (blue arrow). Calculations assume that morphogen is produced by a stripe of cells (from -20 to 0 µm) within an effectively two-dimensional epithelium, so that results may be displayed as one-dimensional graphs. The expression used to calculate these profiles was obtained from Green's function for free diffusion by integrating over space and time (from 0 to 200 s). Diffusion coefficients for the two cases were 5 x 10~7 cm2 s-1 and 10-7cm2s-1. A very similar effect of diffusivity on gradient shape may also be observed for steady state gradients, depending on the conditions that produce the steady state (unpublished observations). (B and C) Simulations of ballistic and diffusive transport, and their responses to barriers. In both cases, 105 moving objects are released-10 per second for 10,000 s-from a single point into a field possessing an impenetrable barrier at one location (arrow). Object positions at the end of the time period are shown by individual dots (upper images), and via histograms of object density along the horizontal axes at the level of the barrier (lower images). In (B) ballistic motion is shown: the objects are tennis balls hit in random directions. Note the large number of balls that accumulate in front of the boundary. In (C) diffusive motion is shown: the objects are morphogen molecules undergoing random walks. Although the slope of the morphogen gradient flattens near the boundary, which may give a visual impression of slight accumulation, the histogram shows that there is no significant build up of molecules at that location.
Figure 1. Consequences of Random, Diffusive Transport
(A) Concentration profiles for a morphogen that is continuously produced at constant rate for a fixed time. The solid and dashed curves display results for two different diffusion coefficients that differ by a factor of 5. For high-threshold morphogen responses (those induced when morphogen levels are above the red line), lower diffusivity shifts the response boundary farther from the morphogen source (red arrow). For low-threshold responses (blue line), the opposite occurs (blue arrow). Calculations assume that morphogen is produced by a stripe of cells (from -20 to 0 µm) within an effectively two-dimensional epithelium, so that results may be displayed as one-dimensional graphs. The expression used to calculate these profiles was obtained from Green's function for free diffusion by integrating over space and time (from 0 to 200 s). Diffusion coefficients for the two cases were 5 x 10~7 cm2 s-1 and 10-7cm2s-1. A very similar effect of diffusivity on gradient shape may also be observed for steady state gradients, depending on the conditions that produce the steady state (unpublished observations). (B and C) Simulations of ballistic and diffusive transport, and their responses to barriers. In both cases, 105 moving objects are released-10 per second for 10,000 s-from a single point into a field possessing an impenetrable barrier at one location (arrow). Object positions at the end of the time period are shown by individual dots (upper images), and via histograms of object density along the horizontal axes at the level of the barrier (lower images). In (B) ballistic motion is shown: the objects are tennis balls hit in random directions. Note the large number of balls that accumulate in front of the boundary. In (C) diffusive motion is shown: the objects are morphogen molecules undergoing random walks. Although the slope of the morphogen gradient flattens near the boundary, which may give a visual impression of slight accumulation, the histogram shows that there is no significant build up of molecules at that location. 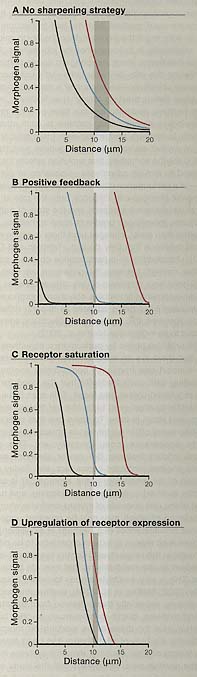 Figure 2. Strategie for Making Sharp Borders (MD) Each panel shows calculated shapes of signaling gradients pro-duced at three rates of morphogen production, over a field of 20 rows of cells, using different sharpening strategies (units of signaling are arbitrary). The shaded box gives the distance from the tenth row to the location where the middle curve falls by half. The width of the box thus reflects the steepness of the curves, whereas the distance between curves reflects robustness to variations in morphogen synthesis rate. The shaded box of panel (A) is extended as a Iight stripe into panels (B)-(D) to facilitate comparison. In (A), no sharpening strategy is shown. The morphogen gradient is a declining exponential, and intracellular signaling is proportional to the amount of extracellular morphogen. In (B), the morphogen gradtant is the same as in panel (A), but there is positive feedback in intracelular siynanig (a key signaling molecule inhibits its own destruction), In (C), morphogen synthesis rates are high enough that receptors are saturated near the start of the gradient. In (D), morphogen stgnafng upregulates expression of morphogen receptors. In (A), (C), and (D), the black, blue, and red curves represent successive 2-fold increases in morphogen synthesis rate, In (B), these curves represent only 8% increases. Notice that in (B), steep declines in signaling over one cell row are easty obtained, but tolerance for variation in morphogen synthesis rate is drarnabcaiy reduced. In (C) and (D), robustness to morphogen synthesis rate is improved, but other performance objectives are compromised (not shown).
Figure 2. Strategie for Making Sharp Borders (MD) Each panel shows calculated shapes of signaling gradients pro-duced at three rates of morphogen production, over a field of 20 rows of cells, using different sharpening strategies (units of signaling are arbitrary). The shaded box gives the distance from the tenth row to the location where the middle curve falls by half. The width of the box thus reflects the steepness of the curves, whereas the distance between curves reflects robustness to variations in morphogen synthesis rate. The shaded box of panel (A) is extended as a Iight stripe into panels (B)-(D) to facilitate comparison. In (A), no sharpening strategy is shown. The morphogen gradient is a declining exponential, and intracellular signaling is proportional to the amount of extracellular morphogen. In (B), the morphogen gradtant is the same as in panel (A), but there is positive feedback in intracelular siynanig (a key signaling molecule inhibits its own destruction), In (C), morphogen synthesis rates are high enough that receptors are saturated near the start of the gradient. In (D), morphogen stgnafng upregulates expression of morphogen receptors. In (A), (C), and (D), the black, blue, and red curves represent successive 2-fold increases in morphogen synthesis rate, In (B), these curves represent only 8% increases. Notice that in (B), steep declines in signaling over one cell row are easty obtained, but tolerance for variation in morphogen synthesis rate is drarnabcaiy reduced. In (C) and (D), robustness to morphogen synthesis rate is improved, but other performance objectives are compromised (not shown).