Молекулярная биология - молодая наука. Открытие, что малые РНК являются центральными в регуляции генов это первый пример важности этой молодой науки. 26 сентября 2008 Albert Lasker Basic Medical Research Award была присуждена пионерам. сделавшими это открытие: Victor Ambros из University of Massachusetts Medical School, Gary Ruvkun из Massachusetts General Hospital and Harvard Medical School и David Baulcombe из University of Cambridge.
Исторически пионеры игнорируются, т.к. значение их открытия понимается спустя годы. Удивительная работа, опубликованная Ambros и Ruvkun, описала первую microRNA и её мишень (in an obscure journal named Cell) игнорировалась поти 10 лет. Между тем, Baulcombe, работал в забытой (at least by US federal funding) области молекулярной биологии растений. Его открытие видов малых РНК, ассоциированных с зависимым от гомологии молчанием, также известным как ко-супрессия или RNA interference, прдзнаменовало выяснение химии и биохимии этих процессов. Работа этих талантливых исследователей фундаменталь но изменила мнение ученых о посттранскрипционной регуляции генов, продемонстрировав широту биологических процессов как на раститальных, так и животных клетках, когда короткие виды РНК играют центральную роль.
Worm Mutants and MicroRNAs
Частично история начиналась с описания двух мутантов у нематоды C. elegans by Chalfie, Horvitz и Sulston в этом журнале в 1981. Эти две мутации, unc-86 и lin-4, изменяли обычно инвариантный прогресс клеточных клонов у круглого червя C. elegans. В частности, различные клеточные клоны у этих мутантов становились "stuck" в определенный момент их развития, повторяя паттерны клеточных делений. В лаб Horvitz laboratory, Victor Ambros заинтересовался этим фенотипом. Базируясь на наблюдении, что мутантные lin-4 черви неспособны откладыать яйца, он предположил, что др. мутации, приводящие к неспособности откладывать яйца, могут иметь сходные аномалии. Проверка мутантов, неспособных откладывать яйца показала, что мутации lin-14, lin-28 и lin-29 были ассоциированы с аномальным прогрессом клеточных клонов (Ambros and Horvitz, 1984). Эти мутации приводили к двум типамнерегулярной прогрессии клеточного клонирования—"precocious" событиям, при которых определенные онтогенетические процессы происходили раньше, чем обычно у животных дикого типа, и "retarded" события, при которых процессы происходили позже, чем обычно у диких животных. По этой причине новые мутации были обозначены как "heterochronic" мутации.
Наиболее интересным геном, открытым в этом исследовании оказался lin-14, различные аллели которого имели противоволожные фенотипы преждевременной или задержанной прогрессии клеточных судеб. Интересно, что полудоминантный задерживающий аллель lin-14 фенокопировал дефекты, наблюдаемые у мутанта lin-4 , тогда как рецессивный преждевременный аллель lin-14 приводил к развитию онтогенетических дефектов, противоположных найденным у мутантов lin-4. Используя 19 индивидуальных мутантных и температуро-чувствительных аллелей lin-14, Ambros вскоре установил, что задерживающие аллели lin-14 являются аллелями с избыточностью функции, тогда как преждевременные аллели lin-14 являются аллелями потери функции и пришел к заключению, что два класса аллелей вызывают свои эффекты в разное время во время развития. Совокупность этих данных показала, что lin-14 действует обычно пониженно на поздних стадиях развития, это приведло к двум сажным вопросам: (1) Как lin-14 регулируется во временив ходе развития и (2) как lin-14 осуществляет своё действие?
Наиболее прямым способом ответить на эти вопросы, стало определние гена lin-14 в молекулярных терминах. К этому времени, Gary Ruvkun, др. сотрудник лаб. Horvitz подключился к исследованиям Ambros. Эти исследования показали, что полудоминантная lin-14 мутация избыточной функции является или делецией и перестройкой 3' области гена lin-14 . Стало ясно, что нарушенная последовательность кодирует элемент, который негативно регулирует активность гена lin-14, но было неясно на каком уровне, ДНК. РНК или белка это происходит. С этого момента Ruvkun и Ambros создали свои собственные группы в MGH/Harvard Medical School и Harvard University, соотв. Два исследователя продолжили свою работу гетерохронных генов червя и в 1989 независимо опубликовали свои работы по этим генам, в частности по lin-14.
Ruvkun получил антитела к продукту гена
lin-14 и продемонстрировал, что этот белок оказывается локализованным в ядре почти всех клеточных клонов, как известно затрагиваемых мутациями
lin-14. Однако экспрессия временно ограничивалась—у животных дикого типа lin-14 присутствует
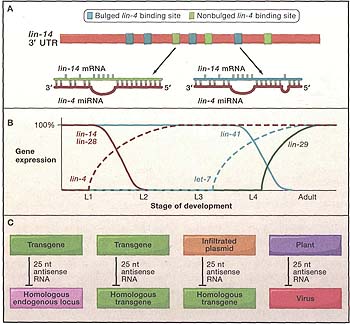 Figure 1. Interactions Revealing the Role of Small RNAs in Gene Regulation
(A) In C. elegans, the lin-4 microRNA is complementary to seven individual binding sites in the 3' UTR of the lin-14 gene. The region of uninterrupted pairing between the 5' end of the microRNA (the heptamer or octamer between bases 1 and 8) and the mRNA is known as the "seed" and is used for microRNA target prediction in animals. Curiously, in the originally defined microRNA/mRNA interaction, the binding sites characterized by "bulged" seed pairing architecture conferred repression on a heterologous UTR, whereas binding sites with uninterrupted seed pairing were less active (Ha et al„ 1996). Figure adapted from Wightman et al. (1993).
(B) The stage-specific expression of heterochronic genes and their interactions as defined by Ambros and Ruvkun in adult worms and in worms at different stages of larval development (L1-L4). (Expression of RNA gene products, dotted lines; protein gene products, solid lines; direct interactions are indicated by like color.) Figure adapted from Reinhart et al. (2000).
(C) The four classes of gene silencing in which Hamilton and Baulcombe initially demonstrated the presence of 25 nucleotide (nt) RNA species that were antisense to the silenced sequences (Hamilton and Baulcombe, 1999).
Figure 1. Interactions Revealing the Role of Small RNAs in Gene Regulation
(A) In C. elegans, the lin-4 microRNA is complementary to seven individual binding sites in the 3' UTR of the lin-14 gene. The region of uninterrupted pairing between the 5' end of the microRNA (the heptamer or octamer between bases 1 and 8) and the mRNA is known as the "seed" and is used for microRNA target prediction in animals. Curiously, in the originally defined microRNA/mRNA interaction, the binding sites characterized by "bulged" seed pairing architecture conferred repression on a heterologous UTR, whereas binding sites with uninterrupted seed pairing were less active (Ha et al„ 1996). Figure adapted from Wightman et al. (1993).
(B) The stage-specific expression of heterochronic genes and their interactions as defined by Ambros and Ruvkun in adult worms and in worms at different stages of larval development (L1-L4). (Expression of RNA gene products, dotted lines; protein gene products, solid lines; direct interactions are indicated by like color.) Figure adapted from Reinhart et al. (2000).
(C) The four classes of gene silencing in which Hamilton and Baulcombe initially demonstrated the presence of 25 nucleotide (nt) RNA species that were antisense to the silenced sequences (Hamilton and Baulcombe, 1999).
в ядрах клеток на самой ранней (L1) стадии личиночного развития, но отсуствует на последующей (L2) стадии развития. Напротив, экспрессия белка у lin-14 мутантов с избыточной функцией сохраняется на всех стадиях личиночного развития и наблюдается даже у взрослых червей. Эти результаты были дополнены генетическими исследованиями Ambros' , предоставившими дополнительные доказательства, что снижение экспрессии белка lin-14 при переходе L1/L2 является критическим для нормального развития. в
Спустя две недели после появления работы Ruvkun's , была опубликована работа Ambros, описывающая анализ эпистаза lin-4, lin-14, lin-28, и lin-29 мутаций. При анализе червей, несущих различные комбинации этих мутаций Ambros расположил гены в виде пути, где ген lin-4 негативно регулирует lin-14 и lin-28. В свою очередь, lin-14 и lin-28 негативно регулируют lin-29, который даже в отсутствии др. трех генов был необходим и достаточен для управления переходом личинок C. elegans во взрослых.
Хотя стало ясно, что ген lin-4 выключает гены lin-14 и lin-28 на личиночной стадии развития, эпистатический анализ не смог однозанчно установить регулирует ли lin-4 оба гена отдельно или один посредством другого. Всё же учитывая сходство фенотипов при
lin-14 избыточности функции и при lin-4 потере функции, оба исследователя предположили, что скорее всего, что lin-4 участвует (вообщето непосредственно) в негативной регуляции lin-14 посредством элемента, делетируемого у мутантов lin-14 с избыточной функцией.
Приблизительно два года спустя Ruvkun опубликовал back-to-back исследование, проливающее дополнительный свет на природу регуляции lin-14. Тщательная молекулярная характеристика генетического локуса, кодирующего lin-14 показала, что мутации lin-14 с избыточной функцией локализуются в 3' untranslated region (UTR) транскриптов, кодируемых геном lin-14, указывая тем самым. что эти мутации не затрагивают белок-кодирующей последовательности. Принимая во внимание, что lin-14 мРНК таких мутантов экспрессируется на нормальном уровне, эти мутации скорее всего нарушают негативный регуляторный элемент, действующий на уровне РНК. в
Путем анализа экспрессии lin-14 белка на гетерохронном мутантном фоне группа Ruvkun продемонстрировала, что lin-14 антагонистически регулируется с помощью lin-28 и lin-4. А именно, в то время как экспрессия белка lin-14 не снижается после стадии L1 у мутантов с потерей функции lin-4, экспрессия белка lin-14 преждевременно исчезает во время стадии L1 на мутантном lin-28 фоне. Эти результаты объяснили предыдущее постороение пути Ambros и показали, что продукт гена
lin-4 (или иногда регулируемый этим продуктом) непосредственно соединяется с областью lin-14 3' UTR, делетируемой у аллелей
lin-14 с избыточной функцией. Осталось прояснить вопросы: каковы молекулярные качественные особенности lin-4, и регулирует ли lin-4 непосредственно или косвенно lin-14? Спустя два года Ambros и Ruvkun опубликовали исследования, ответившие на эти вопросы.
В работе Ambros описывалось молекулярное клонирование гена
lin-4 (Lee et al., 1993). Удивительно, multi-kilobase геномное повреждение, продуцирующее lin-4 фенотип, могло быть комплементровано всего лишь с помощью 693 пар оснований геномной последовательности. Соотв. регионы от
C. briggsae, C. remenei и C. vulgaris также восстанавливали lin-4 фенотип. Однако, когда детальный анализ выявил два блока консервации последовательностей ДНК у 4-х видов нематод, Ambros и его группа не смогли идентифицировать какдидата на роль белок-кодирующей последовательности или канонические старт и стоп кодоны в этих областях. В самом деле, повреждения, призванные нарушать потенциальную кодирующую последовательность не оказывали влияния на восстановление lin-4.
Заинтригованные, авт. продвигались вперед. Northern анализ выявил, что локус
lin-4 продуцирует два обнаружимых вида РНК — минорный в 61 нуклеотида продукт и величиной в -21 нуклеотида продукт. Два продукта были связаны, меньший из продуктов соответствовал наиболее 5' 21 нуклеотидам крупного. Чтобы продемонстрировать, что эти два вида на самом деле влияют на функцию гена
lin-4 в лаб. Ambros скринировали свыше 20,000 хромосом. чтобы идентифицировать вторую мутацию
lin-4, обозначенную Iin-4(ma161). Этот мутантный аллель результат замены цитозина на тимидин, изменеющей позицию 5 двух генных продуктов гена
lin-4. Следовательно, один или оба из этих продуктов может участвовать в обеспечении функции гена
lin-4.
В работе Ruvkun с сотр. продолжились их предыдущие наблюдения, что мутации с избыточной функцией
lin-14 локализуются на 3' UTR в lin-14. Immunoblot и RNase protection эксперименты на червях разных стадий развития однозначно продемонстрировали, что ген
lin-14 регулируется посттранскрипционно стадио-специфическим образом (Figure 1; Wightman et al., 1993). У червей дикого типа хотя белок lin-14 в основном отсутствует после первой стадии личиночного развития, lin-14 мРНК стабильно экспрессируется на всех личиночных и даже взрослой стадии. Используя репортерные трансгены
C. elegans и C. briggsae, группа Ruvkun's продемонстрировала, что 3' UTR lin-14 достаточно для обеспечения наблюдаемой посттранскрипционной регуляции, и что эта регуляция нарушена у двух lin-4 мутантов — как классического аллеля Iin-4(e912), так и вновь изолированного Iin-4(ma161) аллеля. Обе группы отметили, что малые РНК, происходящие с локуса
lin-4 были комплементарны 7 частично повторяющимся последоваетельностям в lin-14 3'UTR, которые делетируются целиком или частично в аллелях lin-14 с избыточной функцией. Ambros и Ruvkun пришли к выводу, что короткие РНК с локуса
lin-4 непосредственно регулируют продукт гена
lin-14 путем спаривания оснований с повторяющимися последовательностями в 3' UTR мРНК lin-14. Т.о., были идентифицированы первая микроРНК и первая мишень для микро РНК.
Группа Ruvkun's по-существу подтвердила модель прямого взаимодействия между lin-4 и lin-14 в последующем исследовании. Они продемонстрировали, что точковые мутации внутри предсказуемых lin-4-связывающих сайтах в lin-14 3' UTR устраняют стадио-специфическую регуляцию репортерных трансгенов
in vivo и что перенос предполагаемых lin-4 сайтов с bulged архитектурой может обеспечить стадио-специфическую регуляцию на гетерологической UTR (Ha et al., 1996). Вскоре после этого группа Ambros's установила lin-28 в качестве второй мишени для lin-4, сгенерировала и охарактеризовала аллель lin-4 с избыточной функцией. В отдельном исследовании было продемонстрировано, что lin-4 действует путем блокирования синтеза белка lin-14 после инициации трансляции lin-14 мРНК (Olsen and Ambros, 1999). Однако, важность этих наблюденийв связи с установленными парадигмами генной регуляции осталась не оцененной. Не было доказательств консервации lin-4 генного продукта помимо
Caenorhabditis, и даже внтри этого рода lin-4 казался едиственным примером малой РНК, которая работает подобным образом.
Всё это изменилось в начале 2000, когда группа Ruvkun's в сотружестве с лабю Horvitz и Ann Rougvie, описала выделение и охарактеризовала ген
let-7 (Figure 1; Reinhart et al., 2000). Подобно lin-14,
let-7 кодирует малую РНК, которая негативно регулирует экспрессию второго гена (lin-41) посредством законсервированных элементов в 3' UTR этого гена. В отличие от lin-14,
let-7 широко законсервирован среди видов животных (Pasquinelli et al., 2000). Спустя 12 мес. после описания let-7 группы Ambros, David Bartel и Thomas Tuschl одновременно опубликовали работы, иллюстрирующие, что существуют сотни генов, подобных lin-4 и let-7, и что это целый класс генов, законесервированных от червей до человека (Lagos-Quintana et al., 2001; Lau et al., 2001; Lee and Ambros, 2001).
Short RNAs and Gene Silencing in Plants
Находка, что существуют сотни генов, кодируют короткие регуляторные РНК в геноме животных была сногшибательной. В начале 1990s, David Baulcombe и др. работали над созданием резистентности к вирусам у растений. Обычной стратегией в то время было активное вмешательство в репликацию вирусов за счет конкуренции за факторы, необходимые вирусам и хозяину посредством избыточной экспрессии трансгенных вирусных белков или структуированных мотивов из нуклеиновых кислот. Однако, вскоре стало ясно, что резистентность к вирусам может быть обеспечена трансгенами даже если трансгены не кодируют белковых последовательностей. Инфекция трансгенных растений вирусами, имеющими гомологичные последовательности приводила к снижению уровней динамического равновесия вирусной и трансгенной РНК, даже если транскрипция с трансгена оставалась неизменной. Эти наблюдения привели к модели, согласно которой резистентность к вирусам обеспечивалась с помощью пост-транскрипционного механизма, который действует. чтобы деградировать вирусную РНК. Сходство между резистентностью к вирусам и зависимым от гомологии молчанием генов у трансгенных растений позволило предположить, что эти два процесса родственны.
Группа Baulcombe's подкрепила взаимоотношения между зависимой от гомологии генной экспрессией и резистентностью к вирусам в серии исследований, опубликованных в сердине 1990s. Впервые была сравнена резистентность к вирусам между несколькими трансгенными линями, и было отмечено, что трансгены, отвечающие за резистентность к вирусам, продуцируют низкие уровни динамического равновесия трансгенной РНК и способны супрессировать при накоплении в транс-положении гомологичных РНК из др. трансгенных локусов (Mueller et al., 1995). Резистентность к вирусам, обеспечиваемая этими трансгенами, была охарактеризована с помощью удивительной специфичности: трансгенные линии, резистентные к определенным штаммам вируса X, не были резистентными к очень близким штамам вирусов.
Второе исследование продемонстрировало, что молчание невирусных трансгенов предупреждает накопление вирусов, видоизмененных, чтобы содержать последовательности, гомологичные с теми трансгенами (English et al., 1996). Было установлено, что идентичность последовательностей скорее, чем происходение последовательностей, диктует взаимодействия между молчащими трансгенными локусами и РНК вирусами у растений. Дополнительные исследования на растениях, которые имеют "recovered" инфекцию линией tomato black ring nepovirus, показали, что эти растения были резистентны к инфекции после вторичной инокуляции вирусом. Хотя выздоровевшие растения оставались чувствительынми к др. типам вирусов, если некодирующая область РНК вируса при первичной инокуляции, оказывалась включенной в гетерологичный вирус, используемый для вторичной инокуляции, растения оказывались резистентными ко второму вирусу. Параллелизм между этой вирусами вызванной защитой и трансгенами индуцированным молчанием гена привело Baulcombe к заключению, что один и тот же механизм, базирующийся на РНК, скорее всего активен в обоих феноменах.
Вскоре после этого группа Baulcombe's использовала Agrobacterium инфильтрацию трансгенного табака, экспрессирующего green fluorescent protein (GFP), чтобы продемонстрировать существование системного сигнала, который может обеспечить сиквенс-специфическое молчание гена трансгенных локусов у растений (Voinnet and Baulcombe, 1997). Системное молчание GFP может быть также индуцировано с помощью локальной бомбардировки ДНК-покрытыми зототыми частицами и может распространяться в нетрансгенные ткани вследствие трансплантации в подвой, в котором были инициировано молчание (Voinnet et al., 1998). Кинетика и паттерн распространения сигнала молчания указывают на то, что он способен распространяться как от клетки к клетке, так и посредством plasmodesmata и систематически посредством phloem; связным, как было предположено, является нуклеиновая кислота.
Все-таки, хотя большинство доказательства подчеркивает, что антисмысловая РНК выступает в качестве медиатора разных форм пост-трансприпционного молчания генов - от косупрессии до антивирусной активности - не было идентифицировано ни одной РНК, которая была бы антисмысловой к targeted последовательностям. В 1996 в обзоре Baulcombe предположил, что методы, используемые, чтобы найти антисмысловую РНК, ассоцированную с пост-транскрипционным молчанием генов, могут пропускать "small or heterodisperse" РНК, которые д. быть скорее всего продуктами кодируемыми растениями РНК-зависимыми РНК полимеразами. Работая с Andrew Hamilton в 1999, он прямо продемонстрировал, что это действительно так.
Проверка трех трансгенных линий табака, обнаруживающих трансгенами-индуцируемое пост-транскрипционное молчание (cosuppression) эндогенного гена выявила смысловую и антисмысловую в ~25 нуклеотидов виды РНК, соответствующие трансгену (Figure 1; Hamilton and Baulcombe, 1999). Поразительно, эти короткие выды РНК отсутствовали в др. растениях табака с тем же самым трансгеном, который не обеспечивал cosuppression. Затем они продемонстрировали, что антисмысловые виды РНК, присутствующие в трансгенных линиях табака, подвергаются пост-транскрипционному молчанию генов, при этом трансгены не обнаруживали какой-либо гомологии с эндогенными последовательностями. Снова виды РНК отсутствовали в линиях табака, где трансгенный локус не замалчивался. Короткие антисмысловые виды РНК, ассоциируемые с молчащими генами, могут обнаруживаться в двух др. моделях молчания генов: во время системного пост-транскрипционного молчания после инфильтрации Agrobacterium и у растений, инфицированных картофельным вирусом X. Стало ясно, что сходство среди разных типов пост-транскрипционного молчания генов, которое отмечается снова и снова, может быть связано с общим знаменателем: короткие виды РНК комплементарны элементу, который замалчивается.
Future Perspectives
The creative studies of Victor Ambros, Gary Ruvkun, and David Baulcombe revealed the importance of small RNAs in the regulation of genes across diverse organisms. The biochemical analysis of RNA interference led to the discovery of small-interfering RNAs (siRNAs), which have become a ubiquitous tool for silencing genes in mammalian cells and hold promise as therapeutic agents to treat disease. MicroRNAs are thought to regulate over half of all mammalian genes, and alterations in their activities are associated with cancer, inflammation, neuronal development, and chronic heart disease. In plants, these RNAs regulate diverse developmental processes by triggering the generation of other frans-acting small RNAs. It is impossible to understand the biology of multicellular organisms without appreciation of the roles that small RNAs play. It is exciting to contemplate that there may be many other types of small RNAs generated by different processes and with different functions. For example, piRNAs are a new class of small RNAs that are prevalent in germ tissue. These may control the expression of repetitive sequences in the germline, at least partially through epigenetic mechanisms. Although many longer noncoding RNAs are known to exist, in most cases their specific functions are yet to be identified. Examples include the relationship of the Xist RNA to inactivation of the X chromosome and of U19 RNA to imprinting of the Igf locus. There are thousands of such RNAs produced in vertebrate systems. The breakthrough research of Ambros, Ruvkun, and Baulcombe underscores the notion that our understanding of the biology of non-coding RNAs has only just begun.
REFERENCES
Ambros, V., and Horvitz, H.R. (1984). Science 226, 409-416.
English, J.J., Mueller, E., and Baulcombe, D.C. (1996). Plant Cell 8, 179-188.
Ha, I., Wightman, B., and Ruvkun, G. (1996). Genes Dev. 10, 3041-3050.
Hamilton, A.J., and Baulcombe, D.C. (1999). Science 286, 950-952.
Lagos-Quintana, M., Rauhut, R., Lendeckel, W., and Tuschl, T. (2001). Science 294, 853-858.
Lau, N.C., Lim, LP., Weinstein, E.G., and Bartel, D.P. (2001). Science 294, 858-862.
Lee, R.C., and Ambros, V. (2001). Science 294, 862-864.
Lee, R.C., Feinbaum, R.L., and Ambros, V. (1993). Cell 75, 843-854.
Mueller, E., Gibert, J., Davenport, G., Brigneti, G., and Baulcombe, D.C. (1995). Plant J. 7, 1001-1013.
Olsen, P.H., and Ambros, V. (1999). Dev. Biol. 276, 671-680.
Pasquinelli, A.E., Reinhart, B.J., Slack, F., Mar-tindale, M.Q., Kuroda, M.I., Mailer, B., Hayward, D.C, Ball, E.E., Degnan, B., Muller, P., et al. (2000). Nature 408, 86-89.
Reinhart, B.J., Slack, F.J., Basson, M., Pasquinelli, A.E., Bettinger, J.C., Rougvie, A.E., Horvitz, H.R., and Ruvkun, G. (2000). Nature 403, 901-906.
Voinnet, O., and Baulcombe, D.C. (1997). Nature 389, 553.
Voinnet, O., Vain, P., Angell, S., and Baulcombe, D.C. (1998). Cell 95, 177-187.
Wightman, B., Ha, I., and Ruvkun, G. (1993). Cell 75, 855-862.
902 Cell 134, September 19, 2008 ©2008 Elsevier Inc.
