Ремоделирование хроматина, обеспечиваемое комплексами Polycomb group (PcG) и trithorax group (trxG), стабилизирует транскрипционные состояния клеток и играет ключевую роль в развитии, плюрипотентности стволовых клеток, и при раке. Исследования на Drosophila указывают на то, что белки PcG действуют. чтобы поддерживать состояние транскрипционного молчания, а белки trxG способствуют экспрессии путем удержания специфических энхансеров доступными (Grimaud et al., 2006; Schwartz and Pirrotta, 2008; Sparmann and van Lohuizen, 2006). Белки PcGи trxG сильно законсервированы между мухами и млекопитающими, но иногда их роли у млекопитающих могут быть противоречть таковым у дрозофилы. Напр., некоторые trxG brahma-родственные белки действуют прежде всего как репрессоры в ES клетках млекопитающих (Ho et al., 2009). У Drosophila большинство известных генов PcGи trxG кодируют ассоциированные с хроматином белки, которые соединяются в виде комплексов со специфическими элементами ДНК, известными как PcG/trxG response elements (PREs). В то время как профилирование по всему геному мишеней PcG показывает, что PREs существуют у млекопитающих, до сих пор PREs были обнаружены только у мух (Boyer et al., 2006; Bracken et al., 2006; Ku et al., 2008; Lee et al., 2006; Marson et al., 2008; Squazzo et al., 2006).
Стержневые компоненты двух основных polycomb комплексов, Polycomb repressive complexes 1 и 2 (PRC1 and PRC2), сильно законсервированы (Levine et al., 2002). PRC2 стержневые субъединицы присутствуют у растений и животных, тогда как таковые в PRC1 присутствуют у мух, C. elegans и позвоночных (Karakuzu et al., 2009). У мух стержневыми субъединицами PRC2 являются Enhancer of zeste (E(z)), Extra sex combs (Esc), Suppressor of Zeste 12 (SUZ12) и NURF-55. Белки E(z) и Esc образуют histone methyltransferase комплекс, который метилирует лизины 27 и 9 гистона H3, создает эпигенетическую метку, которая может быть связана с помощью Polycomb (Pc) белков у мух и мышей (Cao et al., 2002; Czermin et al., 2002; Muller et al., 2002). Pc является стержневым компонентом PRC1, который также содержит polyhomeotic (Ph), Posterior sex combs (Psc) и dRing. Эти наблюдения привели к модели "hierarchical recruitment", которая предполагает, что PcG-зависимое молчание зависит от последовательного действия сначала PRC2 , а затем PRC1 (Wang et al., 2004).
PRC1 и PRC2 стержневые комплексы не соединяются с ДНК непосредственно, а посредством pleiohomeotic (Pho) PcG белка, его гомолога YinYangl (YY1) у позвоночных и родственных Pho-подобных белков связывает специфический мотив ДНК (Atchison et al., 2003; Brown et al., 1998) и может взаимодействовать с субъединицами PRC1 (Wang et al., 2004). Хотя многие PREs содержат Pho-связывающие сайты, этот единственный не может привязывать PcG белки к ДНК in vivo (Brown et al., 2003), и вообще, не существует одинаковых потребностей с последовательностях для PREs.
Первые идентифицированные PREs располагались у мух на комплексах homeotic segment selector (Hox) генов (Chan et al., 1994; Simon et al., 1993). У мышей PcG и trxG белки необходимы для формирования осевого паттерна и для дифференцировки и пролиферации клеток. У Bmi1 и Ringlb (Rnf2) мутантов пролиферативный дефицит может быть устранен с помощью замалчивания реакции на повреждения ДНК Chk2 или генов опухолевого супрессора Ink4a/Arf, соотв. (Liu et al., 2009; Voncken et al., 2003). Однако дефекты формирования осевого паттерна не устраняются и поэтому ключевые онтогенетические мишени для PcG остаются неизвестными. Многие мутации мышей в PcG и trxG генах затрагивают сегментацию заднего мозга эмбрионов, который временно подразделяется вдоль передне-задней оси на 8 компартментов, известных как ромбомеры (r1-r8) (Lumsden and Krumlauf, 1996). Многие паттерны генной экспрессии, включая экспрессию Hox генов, ограничены специфическими ромбомерами. Трансплантационные эксперименты показали, что клетки из соседних ромбомерных доменов не могут перемешиваться но сортируют сами себя в соответствии с характеристиками ромбомеров и профилями транскрипции (Guthrie and Lumsden, 1991), позволяя прямое и легко видимое наблюдение транскрипционных состояний у клетки за клетокой.
У мышей и мух мутации в генах PcG часто сдвигают экспрессию генов сегментации кпереди. Рентгеновскими лучами индуцированная kreisler (kr) инверсия, которая не нарушает кодирующих последовательностей или r5/r6-специфического S5 энхансера гена сегментации MafB/Kreisler (Cordes and Barsh, 1994; Kim et al., 2005), сдвигает экспрессию MafB кпереди (Giudicelli et al., 2003; Sadl et al., 2003): экспрессиия MafB теряется в r5 и гб, но эктопически присутствует на низких уровнях в r3 у kr/+ и kr/kr эмбрионов (Giudicelli et al., 2003; Sadl et al., 2003). Неправильная экспрессия MafB вызывается с помощью эффекта цис-действующего регуляторного элемента(ов), тогда как экспрессия MafB не нарушена у тяжелого гипоморфного аллеля krenu (Sadl et al., 2003).
Мы полагаем, что инверсия kr может anteriorize экспрессию MafB влияя на PRE. Анализ аллеля kr и эффектов хроматин-ремоделирующих агентов и M33 дозы polycomb гена на экспрессию MafB у нормальных и kr/kr эмбрионов, методы изучения PcG- и trxG-зависимой активности PRE у Drosophila, анализ трансгенных мышей и метод иммунопреципитации хроматина в клетках F9 эмбриональной карциномы позволили идентифицировать и оценить bona fide PRE млекопитающих, обозначенных как PRE-kr, которые регулируют ромбомер-специфичную экспрессию MafB .
DISCUSSION
До этого исследования PREs были идентифицированы только у Drosophila. Здесь мы оценили PRE позвоночных, мышиный PRE-kr.
Is Positional Information Encoded within PREs?
Наш анализ инверсии kr указывает на то, что PRE-kr управляет PcG-protein-зависимой репрессией в передней части заднего мозга и trxG-protein-зависимой активацией задней части заднего мозга (Рис. 6E). Комбинация факторов может влиять на такую специфичную для положения функцию. Во-первых. анализ у Drosophila показал, что PREs крепко ассоциируют с ближайшими к ним промоторами (Lanzuolo et al., 2007). Т.о., PRE-kr может взаимодействовать с регуляторными элементами или с ближайшими промоторами Nnat или MafB. Во-вторых, PRE-krфункция может зависеть от состава PcG белковых комплексов, с которыми соединяется. Различные исследования подтвердили избирательные взаимодействия PREs со специфическими субъединицами PcG. Напр., перекрывание M33/Cbx2 со своими гомологами Cbx4, 6, 7 и 8 может вносить вклад в варьирующие эффекты M33 дозы на MafB, члены семейства Cbx/Polycomb также выполнят разные роли в управлении клеточным циклом. Cbx4 не влияет на репликативное старение фибробластов, комплекс же Cbx8-Bmi1 связывает INK4a-ARF локус с преодолением старения, а Cbx7-обеспечиваемое уклонение от старения является Bmi1 независимым (Dietrich et al., 2007; Gil et al., 2004). В-третьих, взаимодействия белков PcG с др. транскрипционными факторами может обеспечивать дополнительную специфичность. Напр., Bmi1 взаимодействует с транскрипционным фактором E2F6, чтобы репрессировать Hoxгены, но действует независимо от E2F6, чтобы репрессировать локус Ink4a-Arf (Courel et al., 2008). Наконец, PRE-kr может быть чувствительным к сигналам, управляющим формированием передне-заднего паттерна, поскольку внешние RARE могут преодолевать PRE-kr-обеспечиваемую репрессию у трансгенных мышей. Отвечает ли PRE-kr избирательно на RA и др. сигнальные пути ещё предстоит установить.
Selective trxG Protein Interactions
In
Drosophila, белки trxG взаимодействуют избирательно с PRE-kr. Лишь trxl/GAGA фактор влияет на PRE-kr-управляемую репрессию у мух. Принимая во внимание, что у позвоночных отсутствуют ортологи Trxl/GAGAF,
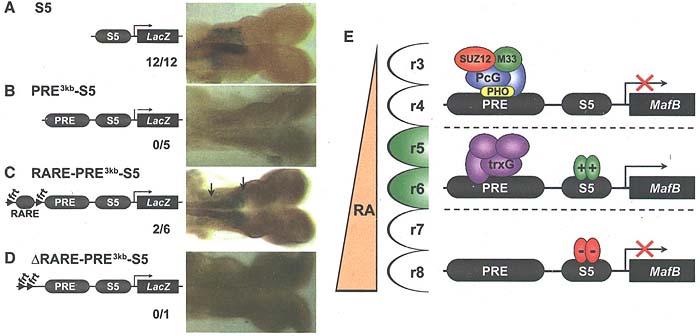 Figure 6. PRE-fcr3kb Silences Induction by S5 in Transgenic Mice
Figure 6. PRE-fcr3kb Silences Induction by S5 in Transgenic Mice
(A) In transgenic embryos, the S5-LacZ (S5) transgene directs LacZ expression in r5/6.
(B) When PRE-to^" is placed upstream of S5 (PRE3kb-S5), LacZ expression is lost.
(C) Addition of Hoxa7 RARE (RARE-PRE3kb-S5) led to recovery of LacZ expression.
(D) Removing the RARE element ARARE-PRE3kb-S5 extinguished LacZ expression in r5/6. Number of transgenic lines with r5/6 expression is shown overtc number of lines analyzed. All embryos are at the 8 somite stage.
(E) A schematic model for the role of PRE-kr in MafB expression. PRC1/2 components act in r3 and r4 at low levels of retinoic acid (RA) to stabilize MafB repr< sion. In r5 and r6, PRE-kr is in an open configuration as a result of retinoic acid signaling, trxG recruitment, and activators, which drive MafB expression via the enhancer. Posterior of r6, unknown inhibitory mechanisms govern S5-dependent repression of MafB.
эффект Trxl на PRE-kr-зависимую репрессию у мух указывает на то, что GAGAF-содержbn trxG комплекс, который также содержит др. белки trxG, законсервирован у мух и мышей и способствует PRE-kr зависимой активации. Компоненты SWI/SNF-related ATP-dependent Brahma хроматин-ремоделирующего комплекса, Brahma и Moira (Crosby et al., 1999; Tamkun et al., 1992), не влияют на PRE-kr-зависимую репрессию у мух. Это возможно отражает избирательное действие SWI/SNF-содержащих комплексов, обнаруживаемое также в нейральном развитии позвоночных (Wu et al., 2007). Дальнейшие исследования необходимы для изучения, может ли PRE-/kr служить в качестве субстрата для специфических SWI/SNF-содержащих комплексов, которые взаимодействуют избирательно с PREs, управляющими ранними нейральными паттерн-формирующими генами.
PREs as Discrete PcG-Binding Platforms
У Drosophila, PcG комплексы соединяются с платформами из дискретных последовательностей (Beisel et al., 2007; Kahn et al., 2006; Papp and Muller, 2006; Schwartz et al., 2006). Сходным образом мы наблюдали разные пики соединения SUZ12 и Bmi1 с PRE-kr в F9 клетках. Напротив, сигнатура H3K27, которая, как полагают накладывается с помощью Ezh1/2-Eed комплекса(ов), покрывает MafBлокус (Czermin et al., 2002; Montgomery et al., 2005). Однако, RA-индуцированное снижение метки H3K27 и потеря связывания PcG оказались высоко локальными. Эти наблюдения указывают на то, что существуют два самостоятельных пула H3K27, лишь один из которых непосредственно зависит от присутствия связывания PRC1 и 2. В согласии с нашими наблюдениями, было предположено, что разные PRC2 комплексы существуют и только один субнабор специфически находит PREs (Nekrasov et al., 2007). Т.о.. др. сигналы, такие как модификации нуклеосом, могут сотрудничать с H3K27 меткой, чтобы пометить и усилить разные SUZ12- и PRC1-связывающе платформы PRE-kr.
Differences in PRC1 and 2 Recruitment
Мы открыли определенные отличия в потребностях последовательностей для связывания PRC1 и 2. Минимальный hcPRE-kr регион может рекрутировать Bmi1, но не SUZ12, связывая эффективно как в экзогенном контексте, таки и в эндогенном контексте. Лишь стабильное связывание PRC1, по-видимому, необходимо для PRE-kr, чтобы репрессировать экспрессию репортерного гена в эктопических местах. Отметим, что у мух рекрутирование PRC1 компонентов PC и PH на PRE трансгенный вставленный сайт служит критерием достоверности эктопически внесенных Drosophilia PREs, но связывание компонентов PRC2 не было изучено подобным образом (Hauenschild et al., 2008 Lyko et al., 1997). Вообще-то PRC2-специфические антитела могли бы выявить отличается ли потребность в стабильной оккупации PRC1 и PRC2 у мух, как это было предложено, исходя из наших находок на F9 клетках. Др. исследования на млекопитающих установили, что PRC1ассоциирует с репрессирующей активностью даже в отсутствие PRC2. CBX8 и Bmi1 обнаруживают сходны уровни связывания со многими генами, даже в отсутствие обнаружимого метилирования H3K27 в Suz12-/- ES rktnjr (Pasini et al., 2007). В Eed нулевых клетках, некоторые компоненты PRC1 рекрутируются на неактивную X (Schoeftner et al., 2006), тогда как матерями предоставляемые компоненты PRC1 обнаруживают Ezh2-независимое поступление на отцовский гетерохроматин (Puschendorf et al., 2008). Более того, PRC1 in vitro способен репрессировать транскрипцию и ингибировать ATP-зависимое ремоделирование хроматина, обеспечиваемое с помощью SWI/SNF комплекса человека - комплекса родственного Drosophila комплексу TrxG Brahma (King et al., 2002).
Комплекс PRC2 может играть др. роль, чем PRC1 в репрессии генов и решении клеточных судеб, как это было подтверждено в исследованиях с такими клетками как Eed нулевые ES клетки (Chamberlain et al., 2008). Наши наблюдения подтверждают, что минимальные последовательности внутри PREs достаточны для стабильного, функционального связывания PRC1, но что рекрутирование дополнительных неизвестных последовательностей подкрепляет стабильность ассоциации PRC2 с PREs.
PRE-kr and the Hierarchical Recruitment Model
Наше наблюдение, что нокдаун SUZ12 затрагивает связывание Bmi1 с эндогенными и эктопическими PRE-kr хорошо согласуется с моделью "иерархического рекрутирования", которая предполагает последовательное действие PRC2 и 1. Принимая во внимание, что связывание SUZ12 с эктопическими hcPRE-kr очень слабое, мы предположили, что временная, нестабильная ассоциация SUZ12 с эктопическим hcPRE-kr достаточна. чтобы стабилизировать связывание PRC1 - вообще-то путем введения необходимых сигнатур метилирования. Наблюдение не прямо подтверждающее модель "hierarchical recruitment" это то, что BmM нокдаун снижает связывание SUZ12 с эндогенными, но не с эктопическими PRE-kr. Это наблюдение может быть объяснено зависимой от последовательностей ролью PRC1 в поддержании стабильного связывания PRC2 или непрямыми эффектами PRC1 на др. компоненты PRC2, напр., путем снижения их уровня. Т.о., возможная взаимозависимость PRC1 и 2 связывания и функционирования всё ещё не установлена для PRE-kr и PREs.
Strategies to Identify Candidate PREs De Novo
PRE-kr не был идентифицирован в исследованиях профилирования всего генома или с помощью биоинформатики. При использовании первой стратегии PcG-связывающие сайты были идентифицированы у позвоночных как располагающиеся вблизи промоторов, тогда как у мух лишь 30% находится вблизи промоторов, а остальные удалены на десятки килобайт. Эти различия могут отражать преимущественное использование недиффернцированных ES клеток позвоночных в противовес псевдо-диффернцированным типам клеток и эмбрионам у Drosophila (Boyer et al., 2006; Bracken et al., 2006; Ku et al., 2008; Lee et al., 2006; Marson et al., 2008; Schuettengruber et al., 2009; Schwartz et al., 2006; Squazzo et al., 2006). На gene-by-gene основании, эти данные трудно интерпретировать. Большинство проделано на ES клетках, где роль PRC2 или PRC1 остается менее ясной. Напр., ES клетки с мутациями в некоторых PRC генах и с отсутствием обнаружимых H3K27me3 не обнаруживали дефектов плюрипотентности (Chamberlain et al., 2008). Существует также несовершенная корреляция с окупацией промотора PRC1/2 и генной экспрессией. В случае MafB, занятие с помощью PRC1/2 PRE-kr не выявляется. Однако, SUZ12 соединяется с промотором MafB в ES клетках человека, но после истощения SUZ12 MafB не активируется (Lee et al., 2006). Т.о., функциональное значение PcG оккупации остается неясным.
Is PRE-kr an Archetypal PRE or a Vertebrate Innovation?
Hindbrain segmentation is a key innovation underlying vertebrate head evolution. Thus, comparative genomic analyses of the PRE-kr sequence might reveal sequence requirements for vertebrate PREs and address whether PRE-kr is a vertebrate innovation or modifcation of an archetypal PRE. The homologous sequences within PRE-kr (Figure 3) are present upstream of all mammalian and the chick MafB orthologs, but not in the genome of zebrafish, in which the hindbrain is segmented but cellularly less complex. BLAST searches (http://genome. jgi-psf.org) did not identify PRE-kr-related sequences in the ceph-alochordate Amphioxus, which is considered to be either among the closest living relatives of craniates or a surviving member of the group from which all chordates evolved (Gee, 2008) and may be hovering at a point in evolution just before hindbrain segmentation emerged (Holland et al., 2008). It remains to be seen to what degree PRE-kr emergence is linked with hindbrain patterning or the need to coordinate fates of complex cell groups.
Concluding Remarks
Our results have wide-ranging implications for PcG mechanisms, as discussed, but also possibily for organization of transcriptional neighborhoods and human disorders.
The finding that the kr inversion translocates a PRE from one rhombomere-specific gene to another might be a coincidence or could suggest that PREs are involved in long-range organization of transcriptional neighborhoods.
With regard to human disease, the self-sorting behavior of rhombomeric cells revealed variable effects of M33 dosage on MafB expression. It is not difficult to imagine that variations in PcG genes or disruptions of PREs perturb neuronal development variably among individuals and contribute to neurobiologic disorders. Indeed, a recent genome-wide association study found linkage between haplotype variants in M33/Cbx2 and schizophrenia (Shi et al., 2009).
We hope that our identification of a vertebrate PRE will guide the identification and validation of other vertebrate PREs and open new doors to mechanistic understanding of PcG-depen-dent silencing.
Cell 138, 885-897, September 4, 2009 ©2009 Elsevier Inc. 893
Сайт создан в системе
uCoz 