Should I Stay or Should I Go: Wnt Signals at the Synapse
 Cell V.130, No.4, P. 593-596, August 24, 2007 | |
|
Several extracellular factors, including Wnt proteins, have been reported to induce synapse formation. In this issue, Klassen and Shen (2007) report that Wnt proteins can also act as antisynaptogenic signals to prevent synapse formation in certain parts of the worm Caenorhabditis elegans. The differential response of axon populations to local Wnt proteins may contribute to the patterning of synaptic connections.
|
Установление функциональных рефлекторных дуг (circuits) в нервной системе нуждается в значительной специфичности при образовании синаптических соединений. Существенный прогресс был достигнут в понимании того, как нейроны, используя позитивные синаптогенные сигналы образуют синапсы на соотв. клетках мишенях. Примерами таких позитивных медиаторов синаптической специфичности являются SYG-1 и SYG-2, открытые у червя C. elegans. Эти две молекулы служат для наведения HSNL моторного нейрона, одного из 4-х нейронов, ответственных за яйцекладки. Без SYG-1 (локализующегося в окружающих эпителиальных клетках, где HSNL нейрон д. сформировать синапс) или SYG-2 (рецептора для SYG-1, расположенного на HSNL аксоне), специфичность синапсов нарушается и формируются эктопические синапсы на несоотв. клетках мишенях (Shen and Bargmann, 2003 and Shen et al., 2004). Сбалансированы ли такие позитивные сигналы негативными сигналами, которые предупреждают образование синапсов в несоотв. зонах мишенях? Klassen and Shen (2007) показали, что Wnt белки могут осуществлять антисинаптогенный эффект, чтобы предупредить иннервацию несоотв. клеток мишеней. Интересно, что Wnt белки, как известно, способствуют образованию синапсов, это говорит о том, что Wnts могут осуществлять про- и антисинаптогенные эффекты на разных клеточных популяциях, влияя на паттерны иннервации.
Klassen and Shen (2007), использовали DA9 нейрон у C. elegans чтобы продемонстрировать, как нейрон может посылать аксон через асинаптическую зону, чтобы достичь своей области мишени, не образуя ненужных синапсов. Нейрон DA9 имеет хорошо закрепленную позицию у C. elegans. Аксон отрастает от тела клетки на вентральной стороне червя и идет в направлении хвоста. Достигнув хвоста аксон поворачивает, переходит на дорсальную сторону и идет кпереди, чтобы сформировать синапс с мышцами дорсальной стенки тела, VD классом ингибирующих нейронов (Figure 1). Хотя мишени для этого нейрона мышцы стенки тела и ингибирующие нейроны находятся на дорсальной стороне червя, он не образует пресинаптических окончаний на наиболее заднем конце аксона. Klassen and Shen (2007) показали, что такое отсутствие образования синапсов является результатом передачи сигналов Wnt, т.о. указывая, что Wnts могут отвечать за распознавание синаптических зон. Wnt Proteins in C. elegans Wnts являются крупным классом сигнальных молекул, участвующих в разнообразных процессах в ходе развития, таких как спецификация осей тела, самообновление стволовых клеток, детерминация клеточных судеб, наведение аксонов и синаптогенез. Известно свыше 16 Wnt белков у позвоночных, все они обладают законсервированным остатком cysteine, который palmitoylated прежде, чем белок будет секретирован.
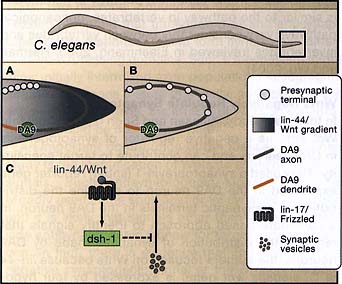 Figure 1. Wnt Activity and Synapse Formation in the Worm
(A) As the axon of the DA9 neuron passes through the tail and through a zone of Wnt expression, presynaptic termini are noticeably absent. However, when the axon passes out of the Wnt expression zone, presynaptic termini are formed.
(B) Mutations in lin-44 (wnt) or lin-17 (frizzled) lead to synapse formation within the asynaptic region of the tail.
(C) A model of how Wnt might prevent synapse formation within the asynaptic zone. Lin-44/Wnt binds to lin-17/frizzled, which signals through dsh-1 to prevent recruitment of synaptic vesicles to the putative active zone.
Figure 1. Wnt Activity and Synapse Formation in the Worm
(A) As the axon of the DA9 neuron passes through the tail and through a zone of Wnt expression, presynaptic termini are noticeably absent. However, when the axon passes out of the Wnt expression zone, presynaptic termini are formed.
(B) Mutations in lin-44 (wnt) or lin-17 (frizzled) lead to synapse formation within the asynaptic region of the tail.
(C) A model of how Wnt might prevent synapse formation within the asynaptic zone. Lin-44/Wnt binds to lin-17/frizzled, which signals through dsh-1 to prevent recruitment of synaptic vesicles to the putative active zone.
Wnts позвоночных соединяются с Frizzled семейством из 7-transmembrane доменовых белков (rev. Miller, 2001). Wnts передают сигналы через несколько разных путей: Wnt canonical путь, планарной клеточной полярности путь и путь кальция. В каноническом пути Wnt соединяется с LRP/Frizzled ко-рецептором, который передает сигналы вниз на Disheveled (Dvl). Dvl затем ингибирует трио белков, GSK3p, Axin и APC. Ингибирование ведет к дефосфорилированию β-catenin, которое запускает его активацию и позволяет ему транслоцироваться в ядро, где он соединяется с TCF/LEF транскрипционными факторами. Неканонические пути у позвоночных также используют передачу сигналов белка Dvl и действуют посредством или JNK или CaMKII и PKC (rev. Ciani and Salinas, 2005). Как и у позвоночных Wnts у C. elegans участвуют в разнообразных процессах, таких как спецификация клеточных судеб и миграция нейробластов. Имеется 5 Wnts (lin-44, egl-20, mom-2, cwn-1 и cwn-2) и 4 Frizzled рецептора (lin-17, mom-5, mig-1 и cfz-2) у C. elegans. Как и у позвоночных имеется канонический и неканонические пути. Канонический путь законсервирован у беспозвоночных и позвоночных с гомологами нематод Dvl (dsh-1, dsh-2, and mig-5), GSK3J3 (gsk-3), Axin (pry-1) и APC (apr-1). Имеется 3 гомолога β-catenin e C. elegans (bar-1, wrm-1 и hmp-2) и только один гомолог TCF/LEF (pop-1). Bar-1 участвует у C. elegans в каноническом пути, где он соед иняется с pop-1, чтобы активировать транскрипцию. Wrm-1, однако, участвует у C. elegans в неканоническом пути и ингибируют pop-1. Хотя канонический путь у нематод сходен с путем у позвоночных, неканонические пути варьируют существенно (rev. Eisenmann, 2005; Herman and Wu, 2004). Wnts Negatively Regulate Synaptogenesis Klassen and Shen (2007) показали, что Wnt действует как негативный регулятор синаптогенеза в нейронах DA9 и может вносить вклад с специфичность мишеней. Они использовали synaptobrevin-1 (snb-1) репортерный ген, экспрессируемый в DA9 нейроне, чтобы проверить распределение пресинаптических окончаний нейрона DA9. В candidate-based подходе для идентификации сигналов, которые регулируют распределение синапсов, образуемых нейроном DA9, авт. сфокусировались на Wnts , т.к. lin-44 (C. elegans wnt ген) экспрессируется четырьмя гиподермальными клетками хвоста, которые создают предполагаемый Wnt градиент в асинаптической зоне. Авт. нашли, что мутации в lin-44 ведут к изменению распределения пресинаптических puncta в ранее асинаптической зоне (Figure 1). Этот фенотип усиливается с помощью мутации др Wnt, egl-20, который экспрессируется на передней и вентральной сторонах хвоста. Мутации в lin-17 (который кодирует Wnt рецептор) воспроизводят этот фенотип. Lin-17 локализуется в асинаптической области аксона DA9, подкрепляя интерпретацию, что локальная передача сигналов Wnt негативно регулирует синаптогенез (Figure 1). Более того, Klassen and Shen (2007) установили, что они могут манипулировать локализацией асинаптической области за счет неправильной экспрессии lin-44 под руководством egl-20 промотора. Они установили, что асинаптическая область расширяется, включая регионы, перекрываемые новым паттерном экспрессии lin-44. Менее удивительными, однако, оказались их находки, что когда они инвертируют Wnt градиент, то только 20% мутантных червей обнаруживают эктопические пресинаптические puncta.
Чтобы понять, как передача сигналов Wnt регулирует образование синапсов, Klassen and Shen (2007) мутировали некоторые компоненты, стоящие ниже lin-17. Они впервые исследовали роль Dvl, ключевого нижестоящего эффектора передачи сигналов Wnt. В согласии с его ролью в передаче сигналов Wnt, они установили, что dsh-1 мутанты имеют повышенные количества пресинаптических puncta внутри ранее асинаптической области. Они также исследовали мутации у ряда компонентов сигнального пути Wnt: двух β-catenin гомологов (bar-1 и wrm-1); TCF/LEF-1 гомолога (pop-1); nematode-like kinase, lit-1 (которая регулирует pop-1); и гомолога axin (pry-1). Ни один из этих мутантов не влиял на распределение синапсов в асинаптических регионах внутри DA9 аксона, указывая тем самым, что новый C. elegans Wnt путь участвует в регуляции образования синапсов. 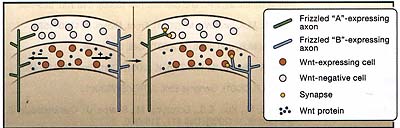 Figure 2. Wnt Signaling and Segregation of Synaptic Inputs
The axons expressing different Wnt receptors (Frizzled "A" and Frizzled "B") are initially intermixed in Wnt-positive and Wnt-negative zones. Frizzled "A" mediates an antisynap-togenic Wnt response, whereas Frizzled "B" mediates a prosynaptogenic response. This leads to segregation of the synapses from two axon populations to the Wnt-negative and Wnt-positive zones. Figure 2. Wnt Signaling and Segregation of Synaptic Inputs
The axons expressing different Wnt receptors (Frizzled "A" and Frizzled "B") are initially intermixed in Wnt-positive and Wnt-negative zones. Frizzled "A" mediates an antisynap-togenic Wnt response, whereas Frizzled "B" mediates a prosynaptogenic response. This leads to segregation of the synapses from two axon populations to the Wnt-negative and Wnt-positive zones. Wnts могут действовать как морфогены (synapto-morphogens), чтобы индуцировать разные исходы исходя из локальной концентрации. Pro- and antisynaptogenic Wnt Signaling Предыдущие исследования на беспозвоночных и позвоночных представили Wnts как позитивные регуляторы синаптогенеза, поэтому находки Klassen and Shen (2007) столь возбуждающие. Исследования на культурах мозжечка мышей показали, что Wnt7a кондиционированная среда увеличивает количество и размер окончаний mossy волокон (Hall et al., 2000). Этот сигнал, как полагают, действует посредством Dvl, который ко-локализуется с пресинаптическими маркерами. Культуры мозжечка. происходящие от мышей, лишенных Dvl, также обнаруживали снижение пресинаптических puncta (Ahmad-Annuar et al., 2006). Исследования in vivo показали снижение размеров пресинаптических окончаний на 10 день постнатального развития, но эти различия не обнаруживались на 15 день (Hall et al., 2000), вообще-то из-за присутствия др. Wnts в мозжечке (Salinas et al., 1994). Это снижение слегка усиливается у мышей с отсутствием Wnt7a и Dvl (Ahmad-Annuar et al., 2006). Хотя эти эксперименты подтверждают возможную позитивную роль Wnts в синаптогенезе, но данные пока ограничены относительно того, могут ли Wnts влиять на количества функциональных синапсов. Patterning of Synaptic Inputs Как Wnts осуществляют как позитивные, так и негативные эффекты на формирование синапсов? Учитывая огромное разнообразие Wnt белков, существует одна возможность, что разные Wnt белки действуют как про- и антисинаптогенные сигналы. Др. интересная возможность в том, что один и тот же Wnt белок может осуществлять про- и антисинаптогенный эффекты в разных популяциях нейронов, которые экспрессируют разные Frizzled рецепторы или нижестоящие белки мишени. Т.о., две популяции нейронов, экспрессирующие разные Frizzled рецепторы, которые передают про- и антисинаптогенные Wnt реакции, д. формировать синапсы в разных Wnt-экспрессирующих доменах. Такой механизм может позволить первоначально перемешанным популяциям аксонов сегрегировать в разные зоны (Figure 2). Наконец. аналогично действию морфогенов, эффекты Wnts на синаптогенез могут быть зависимыми от дозы. |