Большинство работ по передаче сигналов Notch во время нейрального развития фокусируется на начале нейрогенеза, когда клетки предшественники становятся постмитотическими и приобретают определенные судьбы нейральных клеток. Напр., на ст. нейрулы у
и рыбок данио усиливается передача сигналов Notch за счет экспрессии конституитивно активированной формы Notch 1, это предупреждает экспрессию генов нейральной дифференцировки и формирование первичных нейронов, тогда как уменьшение передачи сигналов Notch за счет интерференции с Delta способствует нейрогенезу (Chitnis et al., 1995; Itoh et al., 2003). Однако, передача сигналов Notch также влияет на размер NE во время гаструляции Xenopus (Coffman et al., 1993). Поскольку Notch экспрессируется по всей нервной пластинке в то же самое время, что и NE транскрипционные факторы (Coffman et al., 1990, 1993), а некоторые из этих факторов расширяют нервную пластинку и ингибируют нейральную дифференцировку (Fig. 1), поэтому мы исследовали играет ли передача сигналов Notch роль в регуляции экспрессии этих генов. Мы установили, что: (1) foxD5 действует выше notch 1; (2) усиление передачи сигналов Notch фенокопирует эффекты
, но не меняет экспрессии NE генов, которые ингибируются с помощью foxD5 (soxD, zic1, zic3, Xiro1-3; и 3) передача сигналов Notch необходима для foxD5-обеспечиваемой экспансии доменов экспрессии
. Эти исследования показали, что : (1) передача сигналов Notch действует иерархически ниже foxD5, чтобы обеспечивать экспрессию субнабора NE транскрипционных факторов; и (2) путь передачи сигналов Notch и путь транскрипции
совместно поддерживают нервную пластинку в недифференцированном состоянии.
Во время эмбриогенеза передача сигналов Notch необходима для поддержания и экспрессии нейральных стволовых клеток и клеток предшественников и она ингибирует нейрогенез путем активации Hes1 и Hes5, гомологов дрозофилийных
hairy и Enchancer of split, которые в свою очередь репрессируют экспрессию bHLH гены нейральной дифференцировки (Jarriault et al., 1995; Kageyama and Nakanishi, 1997; Lai 2004; Chiba, 2006; Louvi and Artavanis-Tsakonas, 2006; Chitnis, 2007). Предыдущие исследования выявили большое количество транскрипционных факторов, которые поддерживают нейральные стволовые клетки и клетки предшественники, но интеграция передачи сигналов Notch с регуляцией этих факторов оставалась в основном неизвестной. Было установлено, что передача сигналов Notch действует ниже foxD5, чтобы воздействовать на субнабор NE транскрипционных факторов, которые способствуют недифференцированному нейральному состоянию. Передача сигналов Notch необходима для расширения доменов экспрессии в нервной пластинке
sox11, sox2 и sox3, и она активирует экспрессию
gem и zic2 в нервной пластинке (Fig. 6).
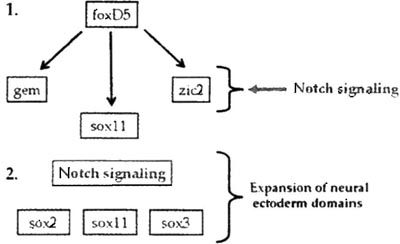 Fig. 6. Notch signaling impacts the expression of NE transcription factors in two ways. (1) foxD5 directly up-regulates the expression of geminin (gem), sox11, and zic2 in both the neural plate and the ventral epidermis. Notch signaling, acting downstream of foxD5, co-regulates gem and zic2 expression levels, but only in the neural plate; it does not up-regulate sox11 expression levels. (2) Downstream of foxD5, Notch signaling is required for the expansion of the neural plate domains of the sox genes.
Fig. 6. Notch signaling impacts the expression of NE transcription factors in two ways. (1) foxD5 directly up-regulates the expression of geminin (gem), sox11, and zic2 in both the neural plate and the ventral epidermis. Notch signaling, acting downstream of foxD5, co-regulates gem and zic2 expression levels, but only in the neural plate; it does not up-regulate sox11 expression levels. (2) Downstream of foxD5, Notch signaling is required for the expansion of the neural plate domains of the sox genes.
Notch Signaling Expands the Neural Plate Domains of Sox Genes Downstream of foxD5
Гены sox2 и sox3, которые принадлежат к подгруппе SoxB1 HMG-box транскрипционных факторов, экспрессирующихся на высоком уровне в NE и в эндогенных нейральных стволовых клетках в ЦНС взрослых. Функционально они поддерживают нейральные стволовые клетки и клетки предшественники и ингибируют нейрональную дифференцировку (Avilion et al., 2003; Bylund et al., 2003; Graham et al., 2003; Ferri et aL, 2004; Pevny and Placzek, 2005; Weg-ner and Stolt, 2005; Wang et al., 2006; Cavallaro et al., 2008; Dee et al., 2008; Kim et al., 2008). Имеются некоторые противоречивые данные относительно того, как передача сигналов Notch и функции генов SoxB1 взаимодействуют. Некоторые сообщения показывают, что Sox2 действует иерархически выше передачи сигналов Notch, поскольку избыточная экспрессия Sox2 позитивно регулирует экспрессию Notch1, Notch лигандов и их нижестоящих генов мишеней (Bani-Yaghoub et al., 2006). Более того, Sox2 может непосредственно регулировать экспрессию гена notch1 путем соединения с его 13th интроном, который обладает биологической функцией в ретинальных нейральных предшественниках (Taranova et aL, 2006). Однако имеются также доказательства, что Sox2 отвечает на передачу сигналов Notch: (1) т.к. мы также выявили здесь, что экспрессия NICD расширяет домен экспрессии sox2 в нервной пластинке (Glavic et al., 2004); и (2) в клетках нервного гребня птиц, sox2 ингибирование дифференцировки находится ниже передачи сигналов Notch (Wakamatsu et al., 2004). Наконец, некоторые исследования показали, что передача сигналов Notch и работа SoxB1 параллельно ингибируют нейрональную дифференцировку (Bylund et al., 2003; Holmberg et al, 2008). Наше исследование обнаруживает дополнительную ступень в пути, показав. что экспансия sox2 и sox3 достигается с помощью foxD5 во время формирования нервной пластинки и нуждается в передаче сигналов Notch (Fig. 6).
Ген sox11, член подгруппы SoxC, также строго экспрессируется в NE и участвует как в нейральной индукции, так и нейральной дифференцировке (Uwanogho et al., 1995; Hargrave et al., 1997; Hyodo-Miura et aL, 2C02; Bergsland et aL, 2006). Хотя имеются доказательства, что sox11 участвует в нейрональной дифференцировке ниже bHLH пронейральных генов, наша предыдущая работа показала, что в эмбриональной NE sox11 функционирует в тандеме с генами
gem и zic2, чтобы расширять в нейральной пластинке экспрессию
sox2 и sox3 (Fig. 1; Yan et al., 2009). К сожалении в предыдущем исследовании не было выявлено взаимодействия между передачей сигналов Notch и sox11. Здесь нами установлено, что передача сигналов Notch необходима для foxD5-обеспечиваемой экспансии экспрессии sox11 в нервной пластинке, но она не усиливает уровни sox11 в NE.
Notch Signaling Up-Regulates Some NE Transcription Factors in Parallel With foxD5
Факторы foxD5, gem и zic2 охарактеризованы также как ингибиторы нейральной дифференцировки. foxD5 расширяет нервную пластинку и ингибирует экспрессию маркеров осевой дифференцировки в NE (en2, Krox20) и нейральной дифференцировки (ngnr1, neuroD, n-tubulin), приводя к интерпретации, что они действуют, чтобы поддерживать NE в незрелом состоянии (Sullivan et al., 2001). Это подтверждается наблюдениями, что foxD5 транскрипционно активирует gem и zic2 и транскрипционное репрессирует NE гены, которые способствуют нейральной дифференцировке (Yan et aL, 2009). Ген gem расширяет нервную пластинку, экспрессируется в нейральных стволовых клетках и противодействует Brg1, белку, который необходим для активации bHLH генов нейральной дифференцировки (Kroll et al., 1998; Seo et al., 2005; Seo and Kroll, 2006; Spella et al., 2007). Ген zic2 расширяет нервную пластинку и экспрессируется в клетках нервных предшественников и ингибирует начало нейрогенеза (Brewster et aL, 1998; Nakata et al., 1998; Aruga, 2004; Mer-zdorf, 2007).
Роль передачи сигналов Notch в экспрессии этих трех NE генов ранее не была исследована. Мы показали, что экспрессия notch 1 находится ниже foxD5. Интересно, что строгая передача сигналов Notch, сопровождаемая экспрессией NICD, активирует gem и zic2 в нейральной эктодерме. Т.о., помимо того, что являются непосредственными транскрипционным мишенями foxD5 (Yan et al., 2009), эти гены, по-видимому, действуют или независимо или ко-регулируются с помощью передачи сигналов Notch. Важно в будущем выяснить эти две возможности. Тот факт, что X-Delta- 1STU также активируют gem и zic2 указывает на то, что эти два гена могут дополнительно регулироваться с помощью др. Notch лиганда, такого как Serrate, или др. взаимодействующих путей. Важно выяснить участие разных Notch лигандов в этом сигнальном пути
(e.g., D'Souza et al., 2008).
In conclusion, elucidating the interactions between Notch signaling and the transcription factors involved in neural differentiation has been very important for understanding how this pathway regulates neuronal fate acquisition. We show that Notch signaling also interfaces with a subset of transcription factors that regulate the expansion of the neural plate and prevent neural differentiation. Further study of the role of Notch signaling in the earliest steps of neural fate acquisition should provide insights into the molecular mechanisms controlling the expansion of neural stem and progenitor cells (Gaulden and Reiter, 2008).
Сайт создан в системе
uCoz