Role of polycomb proteins Ring1A and Ring1B in the epigenetic regulation of gene expression
 Int. J. Dev. Biol. 53: 355-370 (2009) doi: 10.1387/ijdb.082690mv | |
|
|
Генерация клеточного разнообразия зависит от эпигенетических регуляторных механизмов. Белки Polycomb group (PcG) являются центральными компонентами эпигенетической регуляции у метазоа. Система, первоначально ассоциированная со стабильностью транскрипционной программы во время развития, используется также в регуляции др. процессов, таких как поддержание плюрипотентности стволовых клеток и пролиферации клеток. Регуляция с помощью PcG осуществляется посредством модификаций хроматина через ковалентые модификации гистонов. Одной из таких модификаций является monoubiquitylation гистона H2A, зависящая от белков Ring1, которая существенны для развития у насекомых и млекопитающих. В эмбриональных стволовых клетках мышей, Ring1A и Ring1B-зависимое убиквилилирование H2A связано с репрессией инициации транскрипции. Исследования клеток млекопитающих выявили многообразие белковых комплексов, содержащих Ring1A и Ring1B, это указывает на широкую регуляторную роль белков Ring1A, Ring1B в эпигенетической регуляции экспрессии генов.
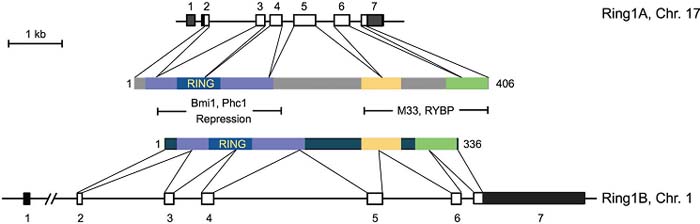 Fig. 1. Genomic structure of the murine Ring1A and Ring1B loci and of the encoded cDNAs. Exons are represented as boxes whose coding parts are empty and 5’ and 3’ UTR are filled. The first intervening sequence in the Ring1B locus, interrupted by slashes, is 22 kb in length and it has be redrawn to the proximity of exon 2. Conserved regions in Ring1A and Ring1B, including the RING fingers, are depicted as colored segments. Proteins binding to the indicated regions, and the repressing domain of Ring1A are indicated. Fig. 1. Genomic structure of the murine Ring1A and Ring1B loci and of the encoded cDNAs. Exons are represented as boxes whose coding parts are empty and 5’ and 3’ UTR are filled. The first intervening sequence in the Ring1B locus, interrupted by slashes, is 22 kb in length and it has be redrawn to the proximity of exon 2. Conserved regions in Ring1A and Ring1B, including the RING fingers, are depicted as colored segments. Proteins binding to the indicated regions, and the repressing domain of Ring1A are indicated.
POLYCOMB PROTEINS 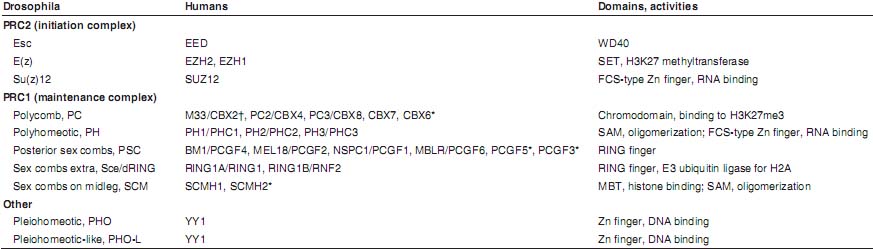 TABLE 2 PROTEINS THAT INTERACT DIRECTLY WITH RING1A AND/OR RING1B 
TABLE 3 PROTEINS IDENTIFIED IN RING1A, RING1B-CONTAINING COMPLEXES 
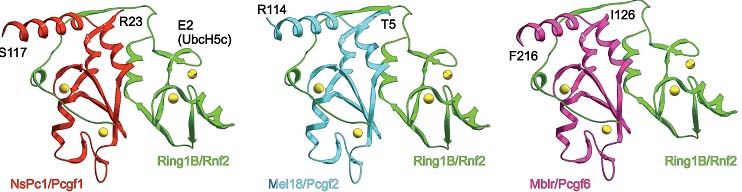 Fig. 3. Models of the associations of murine Ring1B and Pcgfs. Three-dimensional models were built using a knowledge-based protein modeling method based on the given pairwise sequence-template alignments with the A-chain of the 2.5-A-resolution X-ray crystallographic structure of human BMI-1-RING1B polycomb complex as the template [PDB code 2h0d, (Li, 2006)]. Nest, a module of the Jackal software package (Petrey, 2003) was used to build the final structures of the models. Computations were carried out using a SGI Octane workstation. Ribbon representation of backbones in green (Ring1B/Rnf2), red (NsPc1/Pcgf1), cyan (Mel18/Pcgf2) and purple (Mblr/Pcgf6). The Zn-atoms are displayed as VdW volumes in yellow. The Ring1B loop important for E2 ligase Ubc5Hc association is shown. Numbers correspond to the amino acids of the various Pcgfs proteins included in the models. Fig. 3. Models of the associations of murine Ring1B and Pcgfs. Three-dimensional models were built using a knowledge-based protein modeling method based on the given pairwise sequence-template alignments with the A-chain of the 2.5-A-resolution X-ray crystallographic structure of human BMI-1-RING1B polycomb complex as the template [PDB code 2h0d, (Li, 2006)]. Nest, a module of the Jackal software package (Petrey, 2003) was used to build the final structures of the models. Computations were carried out using a SGI Octane workstation. Ribbon representation of backbones in green (Ring1B/Rnf2), red (NsPc1/Pcgf1), cyan (Mel18/Pcgf2) and purple (Mblr/Pcgf6). The Zn-atoms are displayed as VdW volumes in yellow. The Ring1B loop important for E2 ligase Ubc5Hc association is shown. Numbers correspond to the amino acids of the various Pcgfs proteins included in the models.
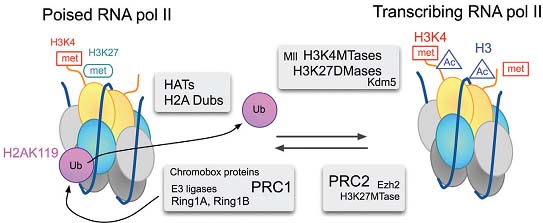 Fig. 4. Model for the reversible switch of transcriptional states at a bivalent promoter. In the inactive state, the E3 ubiquitin ligase acitivity of Ring1A and Ring1B in PRC1 is needed for the maintenance of RNA pol II in a stalled, non-elongating, state. H3K27me3 marks, that depend on PRC2 contribute to stabilize PRC1 association. Recruitment of transcription activating complexes, perhaps through transcription factors bound to DNA sites uncovered by chromatin remodelers, involve coordinated histone modifying activities. Histone acetylation and H2A deubiquitylation, may be trigger modifications of RNA pol II. Demethylation of H3K27 and further methylation of H3K4 would help recruiting TFIID (Vermeulen, 2007) and other componentes of the transcription initiation machinery, thus sustaining transcription. Fig. 4. Model for the reversible switch of transcriptional states at a bivalent promoter. In the inactive state, the E3 ubiquitin ligase acitivity of Ring1A and Ring1B in PRC1 is needed for the maintenance of RNA pol II in a stalled, non-elongating, state. H3K27me3 marks, that depend on PRC2 contribute to stabilize PRC1 association. Recruitment of transcription activating complexes, perhaps through transcription factors bound to DNA sites uncovered by chromatin remodelers, involve coordinated histone modifying activities. Histone acetylation and H2A deubiquitylation, may be trigger modifications of RNA pol II. Demethylation of H3K27 and further methylation of H3K4 would help recruiting TFIID (Vermeulen, 2007) and other componentes of the transcription initiation machinery, thus sustaining transcription.
Perspectives Активность E3 protein ligase, вносящая вклад в поддержание остановившейся (stalled) RNA pol II на репрессированных промоторах, лучше всего характеризует функцию Ring1 белков. Однако остается ещё много неизведанного в отношении специфической роли этой гистоновой модификации и tits зависимости от Ring1A и Ring1B белков: Напр., может ли убиквитилирование H2A, служащее контролю транскрипции, быть отделено от глобального геномного убиквитилирования. Выяснение механизмов доставки, которые рекрутируют Ring1A-Ring1B-содержащие комплексы, также важно. Это может пролить свет на дифференциальное использование Ring1A-Ring1B или др. E3 ubiquitin ligases для достижения убиквитилирования H2A, вобще-то зависящее от стадии клеточной дифференцировки. Получение геномных карт меток H2AUb и их сравнение с таковыми др. гистоновых меток позволит заполнить существенную брешь во взаимоотношениях между разными гистоновыми модификациями и повзолит прояснить рекрутирование и дифференциальное использование E3 ligases на специфических локусах. Регуляция самого процесса ubiquitylation в физиологическом контексте исследована лишь едва. Наконец, дальнейшее исследование связи между H2AUb и RNA pol II stalling, вместе с удалением H2AUb меток может помочь расшифровать молекулярные механизмы роли Ring1A-Ring1B в инициации транскрипции.
Всегда ли Ring1A-Ring1B действует как E3 ubiquitin ligases для H2A в различных комплексах, к которым они прендалежат, неизвестно. Белки Ring1A-Ring1B могут также участвовать в др. слабо охарактеризованных функциях. Напр., хорошо изучена возможность, что Ring1A-Ring1B участвует в сборке и динамике мультипротеиновых комплексов, как показывает зависимость стабильности PRC1 от Ring1B. В этой связи остается неопределенным, каковы возможные функциональные различия между Ring1A и Ring1B.
Изучение функции генов PcG пррошло долгий путь от их первоначальной ассоциации с контролем онтогенетических процессов,регулируемых Hox белками. Надавние успехи показывают, что продуктц генов PcG участвуют в широком круге регуляторных механизмов. Ответы на поставленные выше вопросы помогут лучше понять вклад системы Polycomb модификаторов хроматина в эпигенетическую регуляцию генной экспрессии.
|