Developmental Fate of the Mammalian Myotome  Dev. Dyn. V. 23: P.2898-2910, 2010 Wiley-Liss, Inc. | |
|
The myotome is a segmented paraxial muscle present in all early vertebrate embryos, which in amniotes disappears in mid-embryogenesis, and is replaced by complex epaxial and hypaxial musculature. Little is known about how this transition occurs, Here, we describe the detailed morphogenesis of the epaxial muscles from the epaxial myotome, in rodent embryos. The results show there is no apoptosis of myotomal fibres during the transition, and that the epaxial muscles arise by translocation, re-orientation, and elongation of the myotomal myocytes followed by cleavage of the myotomal masses. Myotomal myocytes transit from a mononucleated to a multinucleated state just before onset of this transformation. Each newly-formed epaxial muscle anlagen includes populations of РахЗ- and Pax7-positive muscle progenitors, with different distributions. Using transgenic mouse embryos bearing a GFP marker for Scleraxis, we show that tendon progenitors are tightly associated with the sides and ends of myotomal myocytes as they re-orient and elongate. Developmental Dynamics 23:2898-2910, 2010 Wiley-Liss, Inc.
|
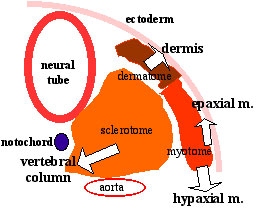
Исследуется способ, с помощью которого epaxial мышцы позвоночных (глубокие мышцы спины) возникают из эмбрионального миотома. Миотом - это метамерная мышца, которая формирутеся внутри сомита, расположен между латеральным дермомиотомом и медиальным склеротомом и является первой мышцей из всех развивающихся у позвоночных. Его моноядерные миоциты занимают всю длину каждого сегмента и располагаются параллельно оси эмбриона (Denetclaw et al., 1997; Kahane et al., 1998; Venters et al., 1999). У птиц и млекопитающих миотомы являются преходящими мышцами и исчезают, чтобы переместиться в epaxial домен став глубокими мышцами спины. Они прикрепляются к позвоночнику и действуют в качестве мышц-разгибателей и получают свою иннервацию от дорсальной веточки смешанного спинального нерва. Взрослые epaxial мышцы представлены тремя основными группами мышц, transversospinalis, longissimus и iliocostalis мышцами, которые располагаются вдоль оси (Vallois, 1922). На торакальном уровне мышца levatores costarum образует четвертую группу мышц, прикрепляющихся к ребрам и поднимающих их во время дыхания (Sato, 1973; Smith and Hol-lydav, 1983). Переход от простого несегментированного миотома к сложной морфологии глубоких мышц спины происходит быстро во время середины эмбриогенеза, но не много известно о том, как это происходит.
 Мышцы спины (второй глубокий слой): Мышцы спины (второй глубокий слой):1 - малая задняя прямая мышца головы; Ключевым вопросом является сохраняются ли исходные миоциты миотома и вносят ли вклад в образование epaxial мышц. Хотя было предположено, что они делают это (Butcher, 1929; Christ et al., 1983; Christ and Ordahl, 1995), но только прямые доказательства сохранения миотомных волокон в дефинитивных аксиальных мышцах, полученные в работе Cinnamon et al. (1999), где hypaxial миотомные миоциты, меченные Dil во время раннего развития, персистировали во всех сегментных межреберных мышцах. Если исходные миоциты сохраняются во всех epaxial и hypaxial производных миотома, то необходимы дальнейшие процессы, которые бы вели к трансформации простого сегментного миотомного каркаса в сложные формы взрослых осевых мышц. Др. возможностями являются те, что волокна миотомов могут подвергаться запрограммированной гибели, а осевые мышцы формируются de novo из предшественников, присутствующих в дермомиотомах или что миотомные волокна могут давать непосредственно некоторые группы мышц с простой сегментной морфологией (напр., межреберные мышцы hypaxially; односегментные transversospinal мышцы epaxially), тогда как мышцы с несегментными формами образуются de novo путем миграции предшественников из дорсо-медиальной губы дермомиотома. Сегодняшнее наше отсутствие понимания судьбы миотома и возникновения взрослых аксиальных мышц резко контрастирует по сравнению с нашим детальным знанием о формировании ранних миотомов (Kalcheim et al., 1999; Venters and Ordahl, 2002; Gros et al., 2004: Holl-way and Currie, 2005) и о возникновении миграторных hypaxial мышц (reviewed in Buckingham, 2001).
Трансформация эмбриональных миотомов в сложный набор epaxial и hypaxial мышц туловища вряд ли осуществляется только благодаря действию миогенных клеток. Мышеная, скелетная и соединительная ткани развиваются в тесной пространственной и временной ассоциации и их морфогенез д. быть синхронизирован так, чтобы формировать функциональные скелетно-мышечные единицы. Stopak and Harris (1982) показали, что соединительно-тканные фибробласты in vitro могут активно организовывать внеклеточный матрикс , так чтобы вызвать сцепление, приводящее к перемещению дисперсных мышечных клеток в организованную мышечную массу в функциональной связи с формирующейся костью. Сходным образомy, Kieny and Chevallier (1979) , а затем Chevallier and Kieny (1982) использовали эксперименты по трансплантациям между перепелом и курицей, чтобы продемонстрировать, что сегрегация и морфогенез мышечных масс конечностей in vivo зависят от соматоплевральной мезенхимы конечностей. В целом, консенсусом экспериментов стало то, что клетки мышечных предшественников необязательно детерминированы в какую-либо специфическую мышцу или регион (Christ et al., 1977; Kardon et al., 2002; Rees et al., 2003); скорее они инструктируются с помощью TCF4-экспрессирующих мезенхимных клеток к дифференцировке в специфическую область и паттерн (Kardon, 1998; Kardon et al., 2003).
Сомитный компартмент, который дает соединительную ткань аксиальных мышц, это синдетом (syndetome) (Schweitzer et al., 2001; Brent et al., 2003). Клетки предшественники из syndetome специфически экспрессируют транскрипционный фактор Sex, но не MyoD (специфичный для клеток мышечных предшественников) или Pax1 (специфичный для склеротомных клеток) (Brent et al., 2003). Позднее Sex экспрессируется в подгруппе развивающейся соединительной ткани, включая сухожилия и апоневрозы (Schweitzer et al., 2001). Недавно клональные исследования показали, что соединительная ткань внутренних мышц mfr;t происходит из Sex-экспрессирующих клеток (Pryce and Schweitzer, unpublished data), поскольку экспрессия ScxGFP трансгенного репортера может обнаруживаться здесь, скорее всего, благодаря персистенции экспрессирующегося на высоком уровне GFP транскрипта и белка. Изучены ранние взаимодействия между миотомом, синдетомом и склеротомом и индукция Sex обнаружила зависимость от FGF белков, секретируемых миотомом (Brent et al., 2003, 2005). Однако мало известно о последних взаимоотношениях между Sex-экспрессирующими клетками и волокнами миотомных мышц в период, когда миотом "трансформируется" в epaxial мускулатуру. Особый вопрос, должны ли Sex-экспрессирующие клетки быть позиционированы соответ. образом, чтобы участвовать в регионализации мышечной ткани в регионе.
После инициального образования эпаксиальных мышц их дальнейший рост и дифференцировка нуждаются в постоянном источнике миогенных предшественников, чтобы увеличивать количество волокон и добавление ядер во время удлинения волокон. Это ставит вопрос об источнике этих предшественников. Клетки эпаксиальных предшественников могут уже присутствовать в зачатке эпаксиальных мышц, т.к. они сегрегируют, чтобы сформировать различные мышечные группы, как это видно из работы Gros et al. (2004, 2005). Однако это никогда не было формально продемонстрировано. Напротив, предшественники могут мигрировать из дорсомедиальной губы дермомиотома, чтобы колонизировать новые эпаксиальные мышечные массы. Эта вторая система должна запускать механизм, сходный с тем, что наблюдается при миграции предшественников конечности.
Целью данного исследования - понять позднее развитие эпаксиальной части миотома млекопитающих и уточнить, как он дает дефинитивные эпаксиальные мышцы. Здесь мы установили, что действительно отсутствует гибель миотомных миоцитов во время этого перехода. Конфокальная микроскопия целых образцов показала, что дифференцирующиеся волокна эпаксиального миотома расщепляются на дискретные массы, одновременно меняя свою ориентацию и длину, чтобы создать зачаток эпаксиальной мышцы. Во время этой транслокации, Sex-экспрессирующие соединительно-тканные клетки тесно ассоциированы с концами и боками миотомных миоцитов, что согласуется с их активной ролью в формообразовании мышц. Когда миотомные волокна распределяются по дискретным группам, которые д. в будущем сформировать эпаксиальную мышцу, то пул РахЗ- и Pax7-позитивных миогенных предшественников смешивается с ними, чтобы обеспечить источник их будущего роста и развития.
|