Переориентация
|
|
A New Spin on Planar Cell PolarityPatricio Olguin and Marek Mlodzik  Cell V. 142, No 5, P. 674-676, 2010 |
|
The generation of planar cell polarity (PCP) and tissue shape during morphogenesis is tightly linked, but it is not clear how. Aigouy et al. (2010) now show in the developing Drosophila wing that PCP initially has a radial orientation that becomes realigned to the proximal-distal axis of organ shape by mechanical forces and cell rearrangements mediated by Dachsous.
REFERENCES
Aigouy, В., Farhadifar, R., Staple, D.B., Sagner, A., Roper, J., Julicher, F„ and Eaton, S. (2010). Cell 142, this issue, 773-786.
Baena-Lopez, L.A., Baonza, A., and Garcia-Bel-lido, A. (2005). Curr. Biol. 15, 1640-1644. Classen, A.K., Anderson, K.I., Marois, E., and Eaton, S. (2005). Dev. Cell 9, 805-817. Harumoto, Т., Ito, M., Shimada, Y., Kobayashi, T.J., Ueda, H.R., Lu, В., and Uemura, T. (2010). Dev. Cell 19. Published online September 2, 2010. 10.1016/j.devcel.2010.08.004.
Lawrence, P.A., Struhl, G., and Casal, J. (2007). Nat. Rev. Genet. 8, 555-563.
Seifert, J.R., and Mlodzik, M. (2007). Nat. Rev. Genet. 8, 126-138.
Shimada, Y., Yonemura, S., Ohkura, H., Strutt, D., and Uemura, T. (2006). Dev. Cell 10, 209-222.
Strutt, D. (2003). Development 130, 4501-4513.
Vinson, C.R., and Adler, P.N. (1987). Nature 329, 549-551.
Wu, J., and Mlodzik, M. (2009). Trends Cell Biol. 19, 295-305.
| |
Большинство тканей и органов состоит из и организовано как слои эпителиальных клеток с добавлением к обычной апикально-базальной полярности эпитилия полярности в плоскости эпителия. Это обзначается как planar cell polarity (PCP). Генетические исследоввания на Drosophila melanogaster установили, что существуют две молекулярные системы, координирующие клетоную асимметрию в плоскости ткани. Сюда входит путь передачи сигналов Frizzled/PCP , содержащий белок Van-Gogh (Vang, также известен как Strabismus/ Stbm) и др. факторы (Strutt, 2003; Seifert and Mlodzik, 2007) и путь, обеспечиваемый посредством protocadherins Fat и Dachsous (Lawrence et al., 2007). Хотя молекулярные взаимоотношения между двумя этими системами неясны, существуют строгие доказательства, что они действуют параллельно, посзожно затрагивая назные эффекторы и могут компенсировать др. др. в некоторых тканях (Lawrence et al., 2007; Wu and Mlodzik, 2009).
В развивающихся крыльях Drosophila клеточные асмимметрии в плоскости эпителия впервые выявляются на ст. поздней куколки вдоль проксимо-дистальной оси (приблизительно 24-30 ч после образования пупария). Главные Frizzled/PCP факторы формируют два самостоятельных комплекса, которые становтся локализованными асимметрично или на проксимальной (Vang/Stbm и ассоциированные белки) или дистальной стороне клеток крыла в куколке (Frizzled и ассоциированные белки). Эти комплексы стабилизируются с помощью взаимодействий типа петель обратной связи между самих себя (Seifert and Mlodzik, 2007; Strutt, 2003). Кроме того, Frizzled и Vang/Stbm белковые комплексы могут быть необходимы значительно раньше для координации глобальной тканевой полярности и PCP в эпителии крыла (Classen et al., 2005; Wu and Mlodzik, 2009).
Подтверждением этой идеи является новое исследование Aigouy et al. (2010)в данном номере ж. Cell, помогающее установить, что субклеточные асимметрии среди белков стержневой группы Frizzled/PCP уже присутствуют на ранних стадиях куколки во время развития крыла (14-15 ч после образования пупария или раньше). Поразительно, но Frizzled/PCP комплексы обнаруживают радиальную полярность, которая перпендикулярна краю крыла (Figure 1A), подтверждая тем самым, что координация глоабальной передачи сигналов Frizzled/PCP устанавливается в крыльях ранних куколок мух. Далее авт. продемонстрировали, что эти ранниее асимметрии на самом деле зависят от передачи сигналов Frizzled-Vang/Stbm, т.к. известно неавтономное поведение мутантных участков клеток frizzled (клонов), влияющее на полярность клеток дикого типа, окружающих мутантные frizzled клетки (Vinson and Adler, 1987) . Поразительно, в противоположность неавтономным эффектам, наблюдаемым на дистальной стороне участков мутантных клеток frizzled в крыльях поздних куколок и взрослых (Vinson and Adler, 1987), ранние frizzled клоны влияли на полярность клеток дикого типа, располагающихся между краем крыла и клоном с осью радиальной полярности. Это подтверждает существование "signaling axis" в направлении края крыла на ранних стадиях. Итак, эти наблюдения Aigouy et al. (2010) показывают, что (1) PCP, обеспечиваемая с помощью передачи сигналов Frizzled-Vang/Stbm, устанавливается во время стадий поздней личинки/ранней куколки по радиальным осям, перпендикулярно краю и что (2) полярность/PCP, наблюдаемая во взрослом крыле является результатом перестроек клеток во время морфогенеза крыла вдоль проксимо-дистальной оси, которые зависят от Dachsous.
Как видоизменяется полярность в проксимо-дистальном направлении в ходе морфогенеза? Поскольку PCP уже установлена на ранних стадиях, то её финальное расположение из радиальной ориентации в проксимо-дистальное направление д. перенаправляться за счёт активной смены положения Frizzled и Vang/Stbm комплексов и/или посредством сдвига или перемещения клеток как целого. Напр., полярность может быть достигнута путем ротаии клеток в направлении дистальной оси, как происходит с ротацией кластеров фоторецепторных клеток (омматидий) в глазах Drosophila в направлении передне-задней оси (Seifert and Mlodzik, 2007). Но как такая ротация может регулироваться? На ранних ст. развития куколки проксимальная половина крылового эпителия (шарнир) и крыловая пластинка одинаковы по рамеру (Figure 1A). Впоследствии, предшествующий и совпадающий с перестройкой PCP, шарнирная область уменьшается и генерирует anisotropic механический стресс на пластинку крыла, который ведет к её удлинению вдоль проксимо-дистальной оси (Figure 1B). Поразительно, элонгация эпителиальных клеток может управлять перестройкой кортикальных микротрубочек вдоль проксимо-дистальной оси, жто, по-видимому, существено для высвобождения Frizzled на дистальной стороне клеток (Shimada et al., 2006). Новая работа, появившаяся в Developmental Cell, Uemura с коллегами (Harumoto et al., 2010) показала, что перед уменьшением шарнирной области расположение кортикальных микротрубочек перпендикулярно краю в проксимальной области пластинки крыла. Это подтверждает генеральную роль ориентации кортикальных микротрубочек для PCP и для перестройки PCP позднее в развитии.
Количественный анализ time-lapse картин крыла куколок (Aigouy et al.,2010) показал, что в ответ на anisotropic стресс, клетки перемещаются относительно др. др. в проксимальном направлении и внутрь с разными скоростями (Figure 1B). Такое поведение взывает сдвиг (shear) и локальную ротацию клеток в основном по часовой стрелке впередней и против часовой стрелки в задней половине крыловой пластинки (Figure 1B). Как результат, PCP переориентируется из радиального направления в проксимо-дистальное внутри крыла (Figure 1С). Интересно, что во время этого процесса ремоделирования глобальная координация и дальнодействующая связанность PCP уменьшается, это может быть причиной того, почему в предыдущих исследованиях PCP во время развития упускали раннюю асимметрию и полярность стержюневых PCP белков. Т.к. клетки крыла куколок вторично упаковываются, поскольку они приобретают гексогональную форму, то глобальная координация и дальнодействующая когерентность PCP снова возрастают, Этот феномен является следствием персистенции Vang/Stbm и Frizzled комплексов на границах, формируемых на ранних стадиях развития и проксимо-дистального выравнивания новых границ, С др. стороны, Frizzled/ PCP стержневые факторы являются необходимыми для гексогональной упаковки клеток, возможно за счет поляризующей мембраны доставки их вдоль прокисимо-дистальной оси (Classen et al., 2005). Т.о., ранняя полярность и клеточная упаковка, д. подпитывать одна др., чтобы задать форму и реполяризовать эпителий.
Было предположено, что система Fat/ Dachsous д. предоставлять "глобальные сигналы", которые ориентируют инициальную полярность Frizzled/PCP комплексов, но эта гипотеза встречает затруднения в виде строгого генетического доказательства, показавшего, что Fat/Dachsous и Frizzled/PCP системы действуют параллельно (Lawrence et al., 2007). Во время личиночной стадии у мух система Fat/Dachsous необходима. чтобы регулировать рост и форму крыла, позднее (по крайней мере частично) путем ориентации оси клеточных делений перпендикулярно краю (Baena-Lopez et al., 2005). Aigouy et al. (2010) показали, что высокая экспрессия Dachsous в шарнирной области не нужна для её контракции, но корректные уровни Dachsous в пластинке крыла необходимы для реакции на anisotropic механический стресс, который ориентирует элонгацию клеток вдоль проксимо-дистальной оси. Более того, дефекты клеточной полярности коррелируют с переворотом ротации локальной ткани в крыле дикого типа с измененными уровнями Dachsous в заднем компартменте, указывая тем самым, что Dachsous насущает клетки способностью отвечать скоординированно на механический стресс. Интересно, Harumoto et al. (2010) показали. что Dachsous и Fat необходимы для выравнивания (align) кортикальных микротрубочек вдоль пркисмо-дистальной оси и что Dachsous оказывает влияние на направлени роста микротрубочек от высокого к низкому уровню Dachsous, подобно его роли в клеточной ориентации. Регулирует ли система Fat/ Dachsous клеточное ремоделирование путем контроля за клеточной полярностью кортикальных микротрубочек или vice versa, предстоит решить.
IВ своем элегантном исследовании, Aigouy et al. (2010) проанализировали временную шкалу событий по установлению PCP в развивающемся крыле
Drosophila. Они предоставили доказательства того, что ранняя Frizzled/PCP core поляризация в направлении края крыла (в радиальной ориентации) перестраивается на поляризацию вдоль поксимо-дистальной оси под действием anisotropic механического стресса и Dachsous-обеспечиваемого ремоделирования ткани. Это заключение согласуется с
Figure 1. Planar Cell Polarity in the Fly Wing
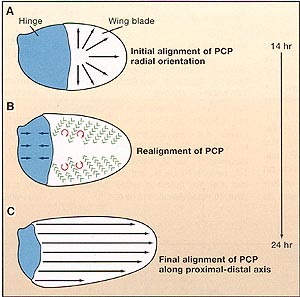 (A) During fly pupal development, the initial axis of planar cell polarity (PCP) is radial, that is, oriented toward the wing margin (black arrows).
(A) During fly pupal development, the initial axis of planar cell polarity (PCP) is radial, that is, oriented toward the wing margin (black arrows).
(B) As development proceeds, the hinge region (blue) contracts creating an anisotropic mechanical stress on the wing blade, resulting in movements of wing cells and realignment of PCP to the proximal-distal axis. Green arrows indicate the direction of cell movement, and red arrows show the direction of cellular rotations. These processes take place between 14 and 24 hr after puparium formation.
(C) The final orientation of PCP is in the proximal-distal axis in late pupal/adult fly wings (black arrows).
и подкрепляется фенотипическими PCP свойствами генов Frizzled/PCP core группы, с одной стороны, и свойствами Fat/ Dachsous системы, с др. стороны. Мухи, несущие мутации в Frizzled/PCP стержневых белках обнаруживают дефекты в PCP по всему крылу. Напротив, система Fat/Dachsous в основном затрагивает полярность в проксимальной половине крыла, эта область сильно зависит от перестройки клеток и ротации во время переключения на проксимо-дистальную PCP ось. Итак, эти наблюдения предоставляют новую основу для понимания генерации PCP и её связи с новыми механизмами, которые формируют органы в целом.
Сайт создан в системе
uCoz