Посещений:  КОНТРОЛЬ ФОРМИРОВАНИЯ ПАТТЕРНА и РОСТА
КОНТРОЛЬ ФОРМИРОВАНИЯ ПАТТЕРНА и РОСТА
Устойчивость, Точность, Масштабируемость
|
|
Pattern, Growth, and Control Arthur D. Lander  Cell 144, March 18, P.955-966, 2011 | DOI 10.1016/j.cell.2011.03.009 |
Systems biology seeks not only to discover the machinery of life but to understand how such machinery is used for control, i.e., for regulation that achieves or maintains a desired, useful end. This sort of goal-directed, engineering-centered approach also has deep historical roots in developmental biology. Not surprisingly, developmental biology is currently enjoying an influx of ideas and methods from systems biology. This Review highlights current efforts to elucidate design principles underlying the engineering objectives of robustness, precision, and scaling as they relate to the developmental control of growth and pattern formation. Examples from vertebrate and invertebrate development are used to illustrate general lessons, including the value of integral feedback in achieving set-point control; the usefulness of self-organizing behavior; the importance of recognizing and appropriately handling noise; and the absence of "free lunch." By illuminating such principles, systems biology is helping to create a functional framework within which to make sense of the mechanistic complexity of organismal development.

Рис.1. | Control Objectives in Morphogenesis
|
Chrysoula Pitsouli and Norbert Perrimon (perrimon@receptor.med.harvard.edu)
The Homeobox Transcription Factor Cut Coordinates Patterning and Growth During Drosophila Airway Remodeling
Sci. Signal., 19 February 2013
Vol. 6, Issue 263, p. ra12
[DOI: 10.1126/scisignal.2003424] | |
Исследовали развитие зачатка, который продуцирует врзослые трахеи у Drosophila, показарно, что гомеобоксны транскрипционный фактор Cut регулирует как рост, так и формирование паттерна, и его эффекты зависят от его концентрации. Определение величины концентрации Cut в резвивающихся предшественниках воздушных путей во время поздней личиночной стадии 3 показало, что клетки развивающихся трахей содержать разные количества Cut, при этом наиболее пролиферирующий регион содержит промежуточные количества Cut, а регион, лишенный Cut обнаруживает дифференцировку. Манипулируя с содержанием Cut, было показано, что Cut функционирует в разных регионах, регулируя пролиферацию или формирование паттерна. Получение профилей транскрипции в популяциях предшественников с разными количествами Cut показало, что передачи сигналов Wingless (известен как Wnt у позвоночных) и Notch являются позитивным и негативным регуляторами экспрессии cut соотв. Более того, был идентифицирован ген, кодирующий рецептор Breathless (Btl, известен как fibroblast growth factor receptor у позвоночных) в качестве мишени транскрипции для Cut. Cut ингибирует экспрессию btl и дифференцировку трахей, чтобы поддерживать развивающиевя клетки воздушных путей в состоянии предшественников. Т.о., Cut участвует в интеграции формирования паттерна и роста в развивающейся эпителиальной ткани.
Идея Waddington's о канализации, что выбираемый нормальный фенотип особенно нечувствителен к генетическим вариациям. В современной литературе термин устойчивость (robustness) является дальнейшим обобщением термина канализация (canalization), включая нечувствительность ко всякого рода пертурбациям средовым и генетическим. Термин точность (Precision) - величина естественной изменчивости в окончательных исходах развития - является мерой устойчивости в отношении природных пертурбаций (напр., существующие генетические вариации, нормальные средовые флюктуации и случайность биохимических процессов).
Частота и степень, с которой эмбриональная регуляция, канализация, устойчивость и точность сталкиваются в развитии, ставит множество вопросов. Существует ли общий принцип, лежащий в основе всех этих феноменов? Существуют ли законсервированные механизмы? Можем ли мы объяснить, как (и почему) разворачиваются такие процессы? Сегодня используются новые методы применительно к биологическим системам с целью выявления лежащих в основе принципов построения дизайна ("design principles"). Обзор посвящен прогрессу нашего понимания онтогенетического контроля и влияния в свете системной биологии. Исследования различных видов животных (Figure 1) обсуждаются ниже; однако необходимо отметить, что подходы системной биологии к морфогенезу растений также сегодня дают (if the pun may be forgiven) существенные плоды (e.g., Jiao and Meyerowitz, 2010; Sahlinetal., 2011).
Complexity, Performance, and Control
Сложная система может быть определена как любая система, в которой достаточно большие количества элементов взаимодействуют простыми способами, чтобы продуцировать неочевидное поведение. Существуют два типа таких систем: те, которые сложны случайно, и те, которые сложны по необходимости. Первые часто изучаются физиками и обычно касаются ситуаций, в которых упорядоченные свойства материи одного уровня описания возникают из коллективного хаоса более низких уровней. Возникновение таких свойств часто суммируется терминами физических законов, подобных Универсальному Закону Идеального Газа или Fick's Законам Диффузии.
Со второго типа сложными системами сталкиваются инженеры, которые проектируют системы, которые бы соответствовали специфическим рабочим характеристикам. Если созданные искусственно системы динамичны (изменяются во времени), а заданные параметры нуждаются в контроле (регулирующее поведение в направлении желательной цели), то количества и типы взаимодействующих компонентов могут быстро достигать точки, при которой поведение системы достаточно непредсказуемым, так что сложная математика или компьютерное моделирование оказываются необходимы для понимания и предсказания их.
Существует явное и сильное сродство между этим вторым типом сложности-обусловленной динамикой и контролем-и картиной, наблюдаемой биологами развития у эмбрионов: динамика, ещё надежное достижение предопределенного завершения. В самом деле, более 60 лет тому назад биологи развития уже предполагали, что "the complex engineering performances of technology are a much more pertinent model of the nature of morphogenesis than are the more elementary phenomena dealt with in basic physics and chemistry" (Weiss, 1950). Хотя в те дни обстоятельства не были подходящими для эксплуатации природного сродства между инженерными разработками и морфогенезом. Как мы увидим, существуют два типа инженерных разработок: forward engineering, которая использует известный набор требуемых рабочих характеристик и строит систему, которая полностью им удовлетворяет и reverse engineering, которая использует знания о том, как система устроена и выводит требуемые рабочие характеристики, которые необходимо воссоздать таким способом. В течение большей части 20-го столетия биологи развития были не готовы воспроизводить любую из них. Сегодня они всё чаще используют оба подхода.
Forward Engineering Pattern
Если мы будем понимать требуемые рабочие характеристики ("performance objectives") в биологии, как соответствующие тому, для чего естественный отбор их выбирает-т.e., тому, что биологи эволюционисты называют приспособленность ("fitness")-тогда мы увидим, что одной из проблем forward engineering является то, что биологи редко обладают полным пониманием того, что вносит вклад в приспособленность (за исключением вообще-то одноклеточных организмов в простых условиях). Более того, даже если требуемые рабочие характеристики, которые движут эволюцию индивидуальной биологической системы, известны, нет гарантии, что forward engineering подход сможет придти к тому же самому решению как в природе. Попросите инженера построить мост и он не будет похож на какие-либо др. мосты. Эта точка зрения иллюстрируется историей паттернов Тьюринга в биологии развития.
Название Turing pattern происходит из плодотворной работы Alan Turing's, которая также внесла понятие морфоген (Turing, 1952). Она описывает решение генеральной проблемы создания повторяющихся паттернов в пространстве. Благодаря дальнейшей разработке работы Тюринга (Gierer and Meinhardt, 1972; Meinhardt and Gierer, 2000), мы знаем, что такие паттерны имеют тенденцию возникать в системе пространственно выстроенных, эквивалентных компонентов (напр., клеток), если они продуцируют как "активирующий", так и "ингибирующий" сигналы, которые распространяются с разными скоростями. В зависимости от деталей может быть достигнуто стационарное состояние, в котором появляются пики и провалы продукции сигналов в виде повторяющихся паттернов из точек или полос (Figure 2A). Turing паттерны являются примером класса механизмов
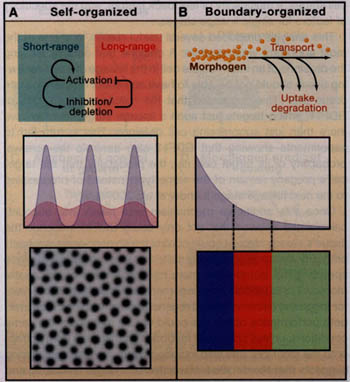 Figure 2. Two Modes of Organization in the Control of Pattern
Figure 2. Two Modes of Organization in the Control of Pattern
The performance objectives of patterning systems include both controlling the locations of events relative to each other and controlling them relative to prespecified landmarks. Turing patterns are one example of self-organizing patterns (A). Repeated patterns form spontaneously and exhibit spacings that depend primarily upon the details of local signal activation, inhibition, and spread, with relatively little influence from events outside the system. In contrast, long-range morphogen gradients typify boundary-driven organization (B). They inform cells of their location relative to fixed landmarks. In both cases, morphogens establish a characteristic "length scale" or "wavelength." In the first case (A), pattern is a direct reflection of that scale, such that elements (spots or stripes) occur once per length scale. In the second case (B), the length scale simply determines how gradually "positional information" decays over space; where pattern elements occur (blue, red, green blocks) depends upon how cells interpret the positional information they receive.
наз. "само-организующими", поскольку локализация и пространственное распределение элементов возникает в результате локальных взаимодействий, а не в результате инструкций передаваемых откуда-то. because the location and spacing of elements emerge out of local interactions, and not through instructions that come from elsewhere. Первоначальные надежды, что Тьюринговские процессы смогут предоставить простое объяснение для всех периодических паттернов развития-от маркирования кожных паттернов морских ракушек до сегментации эмбрионов-не реализовались. В особенности в отношении ранних, высоко точных событий, таких как спецификация эмбриональных сегментов, за 30 лет интенсивная экспериментальная генетика не смогла воссоздать простые диффузионные пары активтор/ингибитор для таких случаев. Вместо этого эти работы обнаружили тенденцию к поддержанию мнения, что паттерн организуется с помощью морфогенов, которые формируют дально-дейстующие градиенты, по которым клетки узнают о своем положении. Такая система организована границами (Figure 2B), означая, что позиционная информация кодируется на одной или нескольких границах с морфогенами пассивно передающими эту информацию поперек поля клеток. Однако интерес к паттернам самоорганизации существенно возрос сегодня частично из-за недавнего доказательства участия процессов Тьюринга в спецификации лево-правосторонней оси, в формировании скелетного паттерна конечностей позвоночных, формирования паттерна эктодермальных органов у млекопитающих и птиц, паттернов пигментации кожи, морфогенеза ветвления в легких и регенерации головы у гидр (reviewed by Kondo and Miura, 2010).
Недавняя работа по паттернам пигментации кожи у рыб (Yamaguchi et al., 2007) оказалась особенно инструктивной, поскольку она использовала преимущества того факта, что самоорганизующиеся механизмы являются прирожденно регулятивными, т.e., они могут локально репарировать сами себя. Более того, точный способ, с помощью которого пигментные полоски отвечают на хирургические манипуляции у рыб, строго характерны для Тьюринговского процесса. Как мы увидим позднее, регулятивная природа самоорганизующихся паттернов может, как помочь, так и препятствовать образованию чёткого паттерна, факт, который может объяснить, почему границами организованные механизмы также необходимы для формирования паттерна.
Работа с формированием паттерна пигментации рыб также подчеркнула тот факт, что создание Тьюринговских процессов не обязательно нуждается в секретируемых, диффундирующих активаторах и ингибиторах (Kondo and Miura, 2010). Тьюринговский процесс является математической абстракцией, которая предполагает продукцию и деструкцию взаимодействующих перемещающихся сигналов. Нет ограничений, накладываемых на молекулярные детали сигналов, как они перемещаются или как они взаимодействуют. Возможно, что настоящее значение Тьюринговских паттернов в развитии недооценены, поскольку биологи слишком сфокусировались на поиске определенных типов молекул скорее, чем на общих принципах построения. Отсюда истекает как сила, так и слабость подхода forward engineering в биологии: оно создает прямой путь к построению принципов, но оно не может сказать нам, как эти принципы исполняются в реальных биологических системах.
Reverse Engineering Growth
Классическим определением reverse engineer является индустриальный шпион, который использует только украденные кальки, цифровые данные, из которых производится конкурентный продукт. В отличие от forward engineering, которая продвигается от требуемых рабочих характеристик к построению, reverse engineering стартует с проекта и поиска, чтобы понять требуемые рабочие характеристики. Чтобы осуществить это инженер д. или использовать предсуществующие знания о принципах построения или использовать моделирование и/или симулирование, чтобы изучить всякого рода поведения системы в надежде распознать рабочие характеристики, что могло быть пригодным или желанным.
Reverse engineering нуждается в обширных знаниях "принципиальной схемы" системы, это является одной из причин, почему возможности создать её были столь редки в биологии вплоть до появления исчерпывающих данных по комплементации методологий, таких как геномика, протеомика, насыщающий мутагенез и прочее. И это только половина причины, почему reverse engineering обнаруживает столь выдающуюся активность в системной биологии. Другая заключается в том, что цель reverse engineering-изучить требуемые рабочие характеристики-заполняется только того типа информацией, которая традиционной молекулярной генетике неподвластна: почему компоненты системы далеки от того, что они делают. Чем более массивна биологическая система, тем более важна способность проникновения в суть.
Среди процессов, к которым системные биологи пытаются применить reverse engineered, являются метаболизм, контроль клеточного цикла, стрессовые реакции и бактериальный хемотаксис (Alon et al., 1999; Csikasz-Nagy et al., 2008; Khammash, 2008; Sauro and Kholodenko, 2004). Первыми явными попытками осуществить reverse engineer сложной онтогенетической системы стала работа Odell's по сети передачи сигналов и генной регуляции, которые устанавливали полярность сегментов у эмбрионов дрозофилы и по передаче сигналов Notch в нейрогенезе насекомых (Meir et al., 2002; von Dassow et al., 2000). В обоих случаях, было предположено, что на построение системы влияет необходимость в устойчивости к параметрам неопределенности и внутренних шумов. За этой работой последовали многие исследования других групп по поиску путей, в которых др. известные механизмы формирования паттерна могут быть также устойчивы (reviewed by Barkai and Shilo, 2009; Eldar et al., 2004; Lander et al., 2009b).
Формирование паттерна это только один из фундаментальных процессов морфогенеза, другим является рост. Т.к. рост часто является следствием пролиферации, то данный Обзор сравнивает контроль роста с контролем пролиферации, осознавая конечно, что пролиферация может осуществляться без роста (напр., у ранних эмбрионов) или со значительной задержкой роста. То что рост находится под тонким контролем подтверждается точностью, наблюдаемой в размерах организмов и их частей. Напр., когда генетическая изменчивость контролируется, то головной мозг взрослых мышей варьирует только в области 5% по размеру и количеству клеток (Williams, 2000). Для билатерально симметричных органов (таких как конечности), лево-правостороння изменчивость в размерах также очень мала (Wolpert, 2010).
Такая точность впечатляет в свете того факта, что пролиферация является экспоненциальным процессом, смесью её ошибок. Не более чем 2% уменьшение в длине клеточного цикла через 30 клеточных циклов вызывает более чем 50% увеличение в размере растущей популяции. Маловероятно, что необходимая точность клеточного цикла, обеспечивающая контроль нормальных органов и размер тела, может быть достигнута без определенного сорта процессов обратной связи. В самом деле, идея, что негативная обратная связь участвует в контроле размера органа, уже давно подтверждена исследованиями по регенерации печени и исследованиями in vitro, показавшими, что многие типы клеток продуцируют субстанции, которые супрессируют их собственную пролиферацию (reviewed by Elgjo and Reichelt, 2004). Работа с такими субстанциями, chalones, велась незначительная вплоть доя конца 1990s, когда было установлено, что мыши, дефицитные по члену скмейства TGF-β GDF8 (myostatin), продуцируют избыток скелетных мышц. GDF8 продуцируется мышцами и действует на мышечные предшественники, тем самым полностью выполняет требования к chalone. Затем, GDF11, близкий гомолог GDF8, как было установлено, обладает аналогичным эффектом в самообновлении нервной ткани, обонятельного эпителия мыши (Wu et al., 2003). Др. молекулы, как недавно было предположено, действуют как chalones в разных тканях (reviewed by Lander et al., 2009a).
Базовая модель chalone-в которой chalones замедляют пролиферацию предшественников, непосредственно связанную с размером органа и ткани-слишком типична для reverse engineering. Необходима действительная принципиальная схема того, как обратная связь исполняется в реальном органе. Прогресс в этом направлении достигнут в исследованиях обонятельного эпителия, где предшественники проходят через определенную клональную стадию и GDF11 действует только на очень специфическую стадию, чтобы повлиять на поведение очевидных временно амплифицирующихся клеток, расположенных между стволовыми клетками и дифференцирующимися нейронами (Wu et al., 2003). Далее было установлено, что activin B, др. член семейства TGF-β, также экспрессируется в обонятельном эпителии и также оказывает негативный эффект на пролиферацию, но действует уникально на стадию стволовых клеток, а не на временно амплифицирующиеся клетки. Reverse engineering этой системы (Lander et al., 2009a) привел к математическому поиску, какие требуемые рабочие характеристики д. потенциально встречаться при разнообразии стадии клеточных предшественников, многообразии факторов обратной связи и специфичности факторов для одиночной клональной стадии.
Этот анализ дал несколько важных результатов и один из наиболее важных оказался негативным: chalone, который действует, чтобы замедлять деления промежуточных клеток в клоне самообновляющейся ткани, должен быть неспособным оказывать какой-либо эффект на устойчивое состояние размера ткани. Это указывает на то, что механизм действия GDF11, который нацелен как раз на такую клональную стадию, должен включать в себя больше, чем просто супрессию клеточных делений. Это в свою очередь привело к экспериментам, показавшими, что GDF11 также контролирует возможность обновления своих клеток мишеней, т.е. возможность, что потомство клеток мишеней будет оставаться того же самого типа вместо перехода в следующую клональную стадию (Lander et al., 2009a).
Как только этот дополнительный механизм был принят во внимание, подсчеты показали, что GDF11 д. влиять не только на устойчивость состояния ткани, он д. контролировать её с почти безупречной жесткостью (robustness). Напр., устойчивое состояние становится прочным в отношении скорости клеточного цикла, инициального количества стволовых клеток и скорости клеточной гибели. Более того, это устройство обратной связи также создает механизм для запуска чрезвычайно быстрой регенерации после повреждения. Однако обе требуемые рабочие характеристики не могут встречаться в одних и тех же условиях, если только не подключена дополнительная обратная связь (в этом случае от activin) со стволовыми клетками. Т.о., reverse engineering указывают на то, что детальные взаимодействия клональности, обратных связей и регуляции самообновления, обнаруживаемые в обонятельном эпителии, составляют систему одновременного мощного контроля размера и быстрой регенерации (Lander et al., 2009b; Lo et al., 2009).
The Value of Integral Feedback
Приблизительно в то же время, когда проводились описанные выше исследования на обонятельном эпителии, две группы независимо пришли к заключению, что контроль с помощью обратной связи количества клеток в гематопоэзе также осуществляется прежде всего за счет регуляции самообновления, т.е. посредством контроля прогрессирования клонов, в противовес контрою скорости клеточного цикла. В одном случае это заключение было подтверждено динамикой регенеративных реакций в ответ на трансплантации костного мозга bone marrow transplantation (Marciniak-Czochra et al., 2009). Др. работа (Kirouac et al., 2009) базировалась на результатах комбинации модельного исследования (системный подход к reverse engineering) и модельной подгонки (использование компьютерных алгоритмов для извлечения значений параметров из данных in vitro и in vivo).
Доказательство, что обратная связь специфически нацелена на самообновление предшественников в системах из множественных элементов, указывает на то, что существуют некоторые общего использования свойства, ассоциированные с этим механизмом. В самом деле, проверка показала, существует прямолинейна реализация стратегии инженерной разработки, известная как интегральный контроль обратной связи. Важно, что интегральный feedback контроль описывает стратегию обратной связи в системе сигналов, которая пропорциональна времени интеграла от различий между системами настоящего поведения и его желательного поведения (Figure 3). Контроль целостности наблюдается и в др. биологических системах (такой как бактериальный хемотаксис) (Figure 3A) и, по-видимому, является в общем существенным всякий раз, когда обратная связь д. поддерживать желаемый исход в точности (контроль заданных значений (set-point control)); это объясняет, почему он может жестко поддерживать самообновление ткани в предустановленном размере. Напротив, регуляция с помощью обратной связи скорости прохождения предшественников через ряд клеточных циклов, то что инженеры называют пропорциональным контролем (регуляция с помощью обратной связи клеточной гибели также определяется
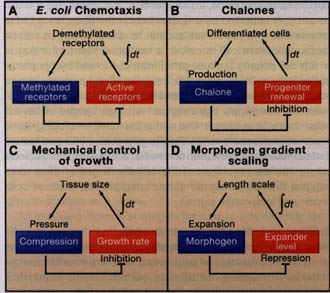 Figure 3. Versatility of Integral Feedback Control
Figure 3. Versatility of Integral Feedback Control
Integral feedback is particularly useful for achieving set-point control, in which a system achieves a prespecified steady-state behavior independent of external (and often many internal) perturbations. The essence of integral control is to feed back a signal that reflects the time integral of error (the difference between the actual and desired states of the system). Biological systems often use this type of control to achieve robust, perfect adaptation, i.e., to return to a zero-activity state even after sustained perturbations. For example, in bacterial chemotaxis (A), integral feedback adaptively modulates signaling to maximize sensitivity to changes in chemoattractant levels (Alon et al" 1999; Yi et al., 2000). Integral feedback in the control of cell growth has been described for two distinct systems. Production of chalones, such as GDF11, by differentiated cells in the olfactory epithelium inhibits progenitor self-renewal (B), providing a feedback signal that increases (decreases) in time as long as the probability of progenitor cell renewal is greater (lesser) than 50% (Lander et al., 2009a). Mechanical compression within the Drosophila wing disc increases with disc size (C), potentially providing a growth inhibitory signal that increases in time as long as cells are proliferating (Shraiman, 2005). Integral feedback can also be used to make a morphogen gradient scale to fit the territory between its source of production and a distant boundary (D). In this case, the morphogen inhibits the production of a molecule that acts at long range to expand the range (length scale) of the morphogen (Ben-Zvi and Barkai, 2010). In such a scenario, buildup of the expander over time provides a time-integrated error signal, which only vanishes when the morphogen gradient expands all the way to the distant boundary.
пропорциональным контролем). Хотя пропорциональный контроль может вызывать некоторую компенсацию отклонений, он в целом не восстанавливает нарушенную систему до заданных значений. Тот же самый интегральный контрольный механизм, с помощью которого достигается надежное поддержание устойчивого состояния в постоянно обновляющихся клонах, таких как обонятельный эпителий или гематопоэз, может также обеспечивать жесткую спецификацию финального размера в необновляющихся тканях, таких как головной мозг. В соответствии с этим, паттерн постепенной экспансии, контракции и исчезновения пула предшественников, который происходит в развивающемся головном мозге, тесно следует ожидаемым следствиям контроля с помощью негативной обратной связи самообновления предшественников (Lander etal.,2009a). В самом деле, измерения вероятностей самообновления предшественников в коре головного мозга показали как раз предсказываемое неуклонное снижение, которое д. продуцироваться контролем с помощью негативной обратной связи (Nowakowski et al., 2002).
В точности, как факторы негативной обратной связи оказываются ответственны за такое поведение в головном мозге, остается неизвестным. Факторы, такие как GDF8, GDF11 и activin присутствуют во многих местах по всей нервной системе. В сетчатке, однако потеря gdfl 1 не вызывает изменений в размере ткани, но характеризуется альтерациями в пропорциях продуцируемых типов нервных клеток, с экспансией некоторых за счет др. (Kim et al., 2005). В нейральной части сетчатки одиночный тип клеточных предшественников, как полагают, дает все дифференцированные клетки, указывая тем самым, что эффекты GDF11's распространяются не только на клетки предшественники, которые самообновляются или дифференцируются, но и также на их выбор, в какой тип клеток будет происходить дифференцировка.
The Range of Control
Несмотря на их возможную важность в регуляции роста и клеточного состава ткани, секретируемые факторы негативной обратной связи могут , в лучшем случае, описывать лишь часть картины. Передача сигналов Notch, напр., также влияет на клеточную пролиферацию и контроль выбора судеб предшественниками (Artavanis-Tsakonas et al., 1999). Посредством латеральной ингибиции Notch может гарантировать, что в точности один тип предшественника сможет возникать внутри определенной области пространства, тип коротко действующего контроля заданных значений. На крайнем противоположном крае действия находятся циркулирующие ингибиторы обратной связи, которые в экспериментах по парабиозу участвовали в контроле за размером печени (Mool-ten and Bucher, 1967).
В самом деле, всякое использование обратной связи для контроля развития обладает характерными пространственными пределами. Напр., секретируемые пептиды ростовых факторов (напр., chalones) , как полагают, действуют внутри эпителиальных тканей в пределах до нескольких сотен микронов, благодаря истощающим эффектам рецепторами опосредованного потребления (e.g., Lander et al., 2009a; Shvartsman et al., 2001). Как могут такие молекулы интегрировать информацию о размерах в значительно более крупной шкале макроскопических органов? Одной из возможностей является то что они действуют на очень ранней стадии развития, когда размеры значительно меньше. По мере роста органа контроль, осуществляемый с помощью обратной связи, д. становиться всё более и более локально автономным. Это д. позволять аккуратную глобальную реакцию на пертурбации, которые затрагивают все места одинаковым образом (напр., генетическую изменчивость, изменения в температуре тела, статусе питания), но не локальные разрушения (напр., физические повреждения части растущей ткани может не вызывать компенсаторного роста в др. месте). Это хорошая основа для размышления о спецификации размера конечностей, которая удивительно точно воссоздает действия частей, которые обладают существенной автономией роста (discussed by Pan, 2007; Wolpert, 2010). Эти наблюдения не говорят о том, что контроль роста достигается без глобальной обратной связи, а о том, что эта глобальна обратная связь просто возникает рано (напр., в зачатках конечностей вместо конечностей). Это делает важной общую точку зрения о точности в развитии: аппарат контроля необходим только в то время, когда соотв. пертурбации обнаруживают тенденцию к возникновению.
Единственное возможное решение контроля роста на крупной пространственной шкале это комбинированные стратегии. Это, по-видимому, происходит в обонятельном эпителии, поскольку размер ткани вдоль апикобазального измерения эпителия (менее 100 µn в толщину) является очень чувствительным к мутациям, которые изменяют экспрессию или функцию GDF11, но латеральная экспансия эпителия в окружающую соединительную ткань (больше многих миллиметров) оказывается менее чувствительной (Kawauchi et al., 2009). Пока же молекулярные механизмы, контролирующие планарную экспансию мало изучены в этой ткани, хотя имеются причины подозревать, что задействована регуляция активности фактора роста фибробластов (Kawauchi et al., 2005).
Одна из тканей, которая стала фокусом значительных экспериментальных исследований, это личиночный крыловой имагинальный диск дрозофилы. Рост крылового диска испытывает влияние со стороны сигналов множественных шкал длины (Edgar, 2006; Martin-Castel-lanos and Edgar, 2002; Nijhout and Grunert, 2010; Schwank and Basler, 2010)-от клетки к клетке, на величину компартмента, на величину диска и гуморальные (гормональные) сигналы. ,Недавние теоретические и экспериментальные работы (Aegerter-Wilmsen et al., 2007,2010; Hufnagel et al., 2007; Nien-haus et al., 2009) предположили, что координация в отношении целого диска может обеспечиваться, по крайней мере частично, с помощью механической обратной связи (длина шкалы которой может быть очень значительной в зависимости от вязкоупругих свойств ткани). Влияние механических эффектов (натяжение, сжимание) на клеточный рост установлено для клеток млекопитающих (Mammoto and Ingber, 2009). В крыловом диске, было установлено, что такие механические эффекты создают благоприятные возможности для интегрального контроля с помощью обратных связей для целого диска (Shrai-man, 2005) (Figure 3C).
Крыловой диск также оказался прекрасным инструментов, чтобы пролить свет на роль пути передачи сигналов Hippo (также известного как Salvador/Warts/Merlin путь) в контроле роста. Молекулярные детали пути Hippo хотя всё ещё появляются и были рассмотрены в обзорах (e.g., Buttitta and Edgar, 2007; Gru-sche et al., 2010b; Haider and Johnson, 2011; Pan, 2007; Reddy and Irvine, 2008) и мы не будем повторяться, но упомянем, что основным источником входящих импульсов в данном пути является протокадерин Fat клеточной поверхности, а главной внутриклеточной мишенью для передачи сигналов Hippo, по-видимому, является рост стимулирующий транскрипционный коактиватор Yorkie (Yki; гомолог YAP позвоночных), который инактивирует передачу сигналов Hippo. В точности, какие критические мишени существуют для Yki, неизвестно, но недавние исследования показали, что подобно GDF11, его функции включают регуляцию самообновления, а не только скорость, с которой клетки проходят клеточный цикл (Haider and Johnson, 2011).
Как и в случае передачи сигналов Notch тот факт, что путь Hippo активируется с помощью лигандов и рецепторов клеточной поверхности (единственным известным лигандом для Fat является Dachsous, также протокадерин клеточной поверхности) это указывает на действие в ранге одной клетки. В самом деле, недавняя работа показала, что путь Fat является центральным в обеспечении хорошо известного in vitro феномена "контактной ингибиции клеточного роста" (Zhao et al., 2007), пример контроля роста в пространственной шкале одиночной клетки. Недавние исследования также установили, что путь Hippo является существенным для компенсаторной пролиферации клеток после повреждения, формы локальной регенеративной реакции. Интересно, что генетические исследования, нацеленные на идентификацию основных детерминант размеров органов, как у позвоночных, так и у беспозвоночных, в основном совпали в отношении пути Hippo-значительно больше, чем пути, контролируемые классическими, диффузными ростовыми факторами (reviewed by Haider and Johnson, 2011; Pan, 2007). Это указывает на то, что пространственные пределы передачи сигналов Hippo могут иногда быть довольно большими. Появилось несколько механизмов того, как это может быть достигнуто.
Во-первых, активация Yki, как было установлено, в средней кишке Drosophila ведет к продукции диффундирующих ростовых факторов и цитокинов, которые стимулируют пролиферацию соседних клетое (Ren et al., 2010; Staley and Irvine, 2010). Во-вторых, очевидно, что Yki может быть сильно активирован с помощью пространственно склонности к оккупированию Fat на одной стороне клетки в противовес другой и что такая склонность может распространяться от клетки к клетке способом, сходным со способом, при котором клеточная полярность передается от клетки к клетке с помощью пути планарной клеточной полярности (при этому пути Fat/Hippo обладают некоторыми общими компонентами) (Reddy and Irvine, 2008). В дополнение к доказательствам что передача сигналов Hippo может действовать поверх границ нескольких клеток, имеются доказательства, что диффундирующие сигналы-включая bone morphogenetic proteins (BMPs) (Rogulja et al., 2008) и факторы, которые активируют Jun киназу (Sun and Irvine, 2011)-также действуют как непосредственные импульсы на регуляцию активности Yki. Итак. эти данные указывают на то, что передача сигналов Hippo может интегрировать и продуцировать коротко- и дальнодействующие сигналы.
Scaling: Matching Pattern to Growth
С самых ранних дней эмбриологии стало ясно, что удивительная устойчивость формирования паттерна прежде всего проявляется на уровне относительного, но не абсолютного, паттерна (т.e., расположение элементов, составляющих паттерн, заложено с учетом размера ткани). Т.о., паттерны, которые возникают у эмбрионов морских звезд, происходящих из изолированных бластомеров, или лягушек, полученных из половинок эмбрионов, обнаруживают нормальные пропорции аномальных размеров.
Необходимость в формировании паттерна, который автоматически соответствует размеру ткани или тела, возникает из того факта, что механизмы контроля роста мощные и адаптивные, т.e., создают постоянные генетические и окружающие условия, они сильно специфицируют точки, задающие размеры, но эти заданные координаты (set-points) сами испытывают влияние со стороны др. факторов (напр., питания, температуры, генетики, и привязки по времени). Прекрасный пример поддающегося адаптации масштабирования (scaling) может быть обнаружен в Dpp градиенте в крыловом диске дрозофилы, который обладает приблизительно константной абсолютной формой в течение всего периода нормального роста диска (Hufnagel et al., 2007), но он сильно меняет шкалу (scales) в ответ на экспериментальные манипуляции, которые увеличивают или уменьшают размер диска (Tele-man and Cohen, 2000). Эти наблюдения указывают на то, что существует точка, задающая масштаб (scaling set-point), но она варьирует в зависимости от стадии развития.
Биологи развития давно пытаются идентифицировать механизмы. ответственные для автоматическое масштабирование (scaling). В самом деле, высокий энтузиазм в отношении оригинальной модели Wolpert's морфогенов как молекул, продуцируемых в источнике и деградирующих на определенном удалении (Wolpert, 1969) был приостановлен автоматическим масштабированием (scaling), которое как таковое достигает определенного порядка (поскольку профиль морфогена в виде прямой линии от источника до устранения (слива); изменения в местоположении слива будет сдвигать все пороговые точки пропорционально). Оказывается, действительно неизвестны градиенты морфогенов, которые бы продуцировались с помощью механизма источник-слив, т.к. необходимо, чтобы морфогены, воспринимаемые клетками, обычно были гарантированы, что они будут деградировать по всему полю его действия скорее, чем только на одном конце. Поскольку такие градиенты не масштабируются автоматически, то были предприняты различные попытки найти стратегии, чтобы сделать их таковыми.
Один из подходов постулировал существование двух морфогенов с противоположных концов поля клеток, при условии, что клетки воспринимают свои позиционные сигналы от соотношения уровней двух молекул. Такая ситуация, которая обычно нуждается в существенной тонкой настройке параметров, была проанализирована для экспоненциально распределенных градиентов морфогенов (McHale et al., 2006) и применена к градиентам более общей формы (Ben-Zvi and Barkai, 2010). Такие модели по существу заменили эффект слива в оригинальной модели Вольперта на независимые позиционные сигналы, которые служат тем же самым причинам. Во всяком случае, масштабирование происходит потому, что поведение этой системы везде оказывается купированным с тем, что происходит на обеих их границах. Такая связь не должна быть прямой. Напр., поскольку скорость пространственного распада морфогена, диффундирующего в эпителии зависит от апикобазальных измерений клеток (которое влияет на скорость протекания морфогена через базальную мембрану), то это будет приводить к тому, что масштабирование градиента морфогена может быть также достигнуто путем сцепления увеличения в планарных изменениях эпителиального слоя с увеличением апикобазальных измерений его клеток (Lander et al., 2011). В самом деле, апикобазальные длины клеток в имагинальных дисках насекомых возрастают параллельно в ростом дисков как целого.
Интригующий класс стратегий по масштабированию недавно идентифицирован как результат попыток reverse engineer спецификации дорсовентральной (D-V) оси у эмбрионов позвоночных. У ранних эмбрионов спецификация клеточных судеб зависит от неуниформности паттерна передачи сигналов BMP вдоль D-V оси. Базируясь на работах с амфибиями и рыбами, а также экстраполируя находки процессов гомологичного формирования паттерна у насекомых, было предположено, что передача сигналов BMP в виде крутого вентро-дорсального градиента, обусловленного комбинированным эффектом инициальной экспрессии BMPs (напр., Bmp 2,4 и 7) на вентральной стороне, а процесс облегченного направленного вентрально транспорта, обеспечивается с помощью BMP-связывающего белка chordin (который продуцируется дорсально, в Шпемановском организаторе). Курьёзно, но дополнительный BMP лиганд, известный как Admp, также продуцируется на дорсальной стороне эмбрионов рыб и лягушек и его экспрессия ингибируется с помощью передачи сигналов BMP. Тот факт, что BMP лиганд экспрессируется на противоположной стороне эмбриона по сравнению с той, где необходима передача сигналов BMP, в комбинации с фактом, что экспрессия этого лиганда чувствительна к сигнальному пути, который меньше всего активен в месте, где он экспрессируется, строго указывает на то, что требуемые рабочие характеристики системы спецификации D-V используют больше, чем только вырабатываемый простой градиент морфогена.
Эта информация привела к открытию, что Admp необходим для масштабирования градиента BMP по отношению к размеру эмбриона (Reversade and De Robertis, 2005). Такое масштабирование бросается в глаза в поведении эмбрионов после хирургических манипуляций, а также в обычной изменчивости между эмбрионами. Позднее была разработана математическая модель для объяснения, как работает такое масштабирование (Ben-Zvi et al., 2008). Недавно был извлечен из этого механизма общий принцип построения, наз. контроль экспансии-репрессии. Этот принцип может быть задействован каждый раз, когда ингибируется передача сигналов градированного морфогена, процесс, который д. иным образом приводить к стремительному росту экспансии самого градиента морфогена (Figure 3D). Если устройство экспансии, расширения (expander) обладает дальнодействующей активностью, то только тогда, когда его экспрессия достигнет почти нуля, градиент морфогена сможет достичь устойчивого состояния. Это будет происходить только, когда градиент морфогена расширяется существенно к краю поля, способного запускать expander, т.e., когда градиент морфогена заполняет ткань, в которой он формирует паттерн. В случае эмбрионов амфибий Admp играет роль expander, и облегчает транспорт посредством действия chordin, гарантируя, что Admp действует на длительном расстоянии.
Подобный механизм масштабирования является примером интегрального контроля с помощью обратной связи (Figure 3D) (Ben-Zvi and Barkai, 2010). Поскольку expander, как предполагается, долгоживущий, то его уровни отражают интеграл времени (time-integral) из ошибок между тем, где градиент морфогена есть, и тем, где он необходим. Только когда ошибка существенно приближается к нулю достигается стабильное состояние. С помощью открытия такой general engineering стратегии в этом механизме авторы пришли к нескольким важным выводам. Во-первых, они получили возможность утверждать, что даже если их детальная, точная модель масштабирования D-V у амфибий (Ben-Zvi et al., 2008) неаккуратна по своей конкретике (как утверждают некоторые; Francois et al., 2009), она, скорее всего, окажется правильной в общем виде. (Процитируем фразу, популярную среди системных биологов, "все модели неправильные; некоторые пригодны." Box, 1979.) Во-вторых, они получили способность перечислить механизмы др. классов, которые математически эквивалентны, даже если механистически непохожи.
Напр., они показали, что расширение градиента может быть обеспечено не только с помощью вторичного морфогена (подобного Admp), но и с помощью любой субстанции, которая регулирует транспорт морфогена, включая диффузионный ингибитор, который также защищает морфоген от отлавливания с помощью рецептора. Интересно, недавно было установлено, что секретируемый белок Pentagone, который негативно регулируется с помощью BMP-родственного морфогена Decapentaplegic (Dpp) в крыловом диске дрозофилы, является мощным expander градиента Dpp (Vuilleumier et al., 2010), это согласуется с ролью в известной способности градиента Dpp менять масштаб в ответ на экспериментальные альтерации размера диска (Teleman and Cohen, 2000). Было также установлено, что секретируемые Frizzled-родственные белки, которые являются конкурентными ингибиторами взаимодействия Wnt-рецептор, действуют как expanders градиентов Wnt (Mii and Taira, 2009) и во время раннего эмбриогенеза амфибий экспрессируются в виде паттернов, согласующихся с негативной регуляцией с помощью Wnts. Хотя необходимо ещё тестировать, действительно ли каждый из этих механизмов по настоящему участвует в масштабировании систем морфогенов, приведенная выше дискуссия иллюстрирует, как систематическое reverse engineering сложных биологических систем может приводить к генерации новых, пригодных для тестирования гипотез. its levels reflect the time-integral of the error between where the morphogen gradient is and where it needs to be
The Management of Noise
Главный вклад системной биологии заключается в повышении осознания роли, которую играют в биологических системах шумы. Здесь шумы определяются как вариации, которые проявляются в случайном или непредсказуемом поведении молекул и клеток. Шумы не являются именно микроскопическими флюктуациями, которые усредняются на макроскопическом уровне; они могут быть как помехой, так и помогать биологической функции. Они могут снижать точность, но они могут также обеспечивать переключение (Hasty et al., 2000), поддерживать и синхронизировать осцилляции (Lewis, 2003), амплифицировать сигналы (Paulsson et al., 2000), или детерминировать динамику стволовых клеток (Hoffmann et al., 2008). Ток шумов через сеть не ведет себя подобно току субстратов через биохимические пути. Шумы могут увеличиваться благодаря стохастическому феномену, помещенному в серию, но могут также уменьшаться благодаря времени интеграции (temporal filtering), а также эффектам обратной и прямой связи, при которых корреляции шумов используются для продукции деструктивной интерференции.
Шумы, которые влияют на формирование паттерна, могут быть как временными, так и пространственными. Эти отличающиеся типы шумов возникают в результате одних и тех же процессов, но последние появляются, когда временные флюктуации являются независимыми клетка от клетки. В общем шумы имеют как временную шкалу (время, в течение которого бывает необходимо усреднение
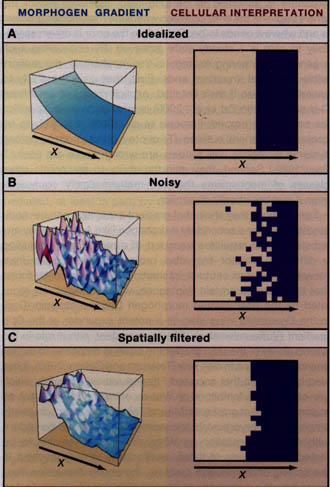 Figure 4. Reducing Noise in Pattern Formation
Figure 4. Reducing Noise in Pattern Formation
In boundary-organized pattern formation, the ability of cells to form organized patterns depends upon the accuracy with which they can measure their positions within a morphogen gradient. Under idealized conditions (A), cells that autonomously adopt a new behavior at a particular threshold value of morphogen concentration will produce a sharp spatial border. In reality, reading a morphogen gradient is fraught with noise: variability in morphogen level, in gene expression, and in cell size, and the stochastic nature of biochemical processes will cause autonomously acting cells to produce "salt-and-pepper" borders (B). Many sources of noise lead to fluctuations on a time scale too slow for cells to compensate simply by integrating signals over time. In principle, processes that enable cells to collaborate with their neighbors can also reduce the noisiness in morphogen gradient interpretation, producing smoother borders. Such collaboration can take many forms. For example, in (C), the noisy signal in panel B was used to drive the production of an activator in a Turing process (the activator induces its own longer-range inhibitor), the level of which was used as a source of positional information. Note the improved border sharpness. Analyses such as this suggest that the combined use of different modes of pattern organization (boundary- versus self-organized) can be useful in achieving robust patterning.
для снижения шумов, в любом данном месте, с помощью данной фракции) и шкалу длин (расстояние, которое д. быть необходимо для усреднения, чтобы снижать шумы в любое данное время с помощью данной фракции). Такое усреднение времени и расстояний не является только функцией амплитуды шумов, но и также их структуры, т.e., будут ли флюктуации происходить независимо во времени (Пуассоновские или короткие шумы ) или в пространстве (отсутствие координации между клетками) или будут обнаруживать корреляции (напр., разрывы или межклеточная кооперация). Одним из величайших сюрпризов в последнее время стало осознание, что большая часть экспрессии генов как у прокариот, так и эукариот является предметом влияния шумов с крупной амплитудой, медленно меняющихся и высоко взрывающихся (Raj etal., 2006). Когда морфогены предоставляют позиционную информацию на большие расстояния, то воздействие шумов ограничивает точность, с которой позиция будет специфицирована (Figure 4). Случайности в диффузии морфогена создают временные шумы в каждом данном месте; это запускает интеграцию времени, в течение которого отвечающие клетки (или ядра, в случает внутриклеточных градиентов морфогена у синцитиальных эмбрионов) д. суммировать свои измерения концентрации морфогена, чтобы адекватно отфильтровать подобные шумы. Для внутриклеточного градиента Bicoid у ранних эмбрионов Drosophila это ограничение может быть существенным поскольку ядра стирают свои "считывания" информации об уровне Bicoid с каждым ядерным делением (которое происходит каждые 10-20 мин). Чтобы бороться с этой проблемой, было предположено, что происходит пространственное усреднение, при этом ядра испытывают влияние от считываний, произведенных их соседями (Gregor et al., 2007). Это иллюстрирует точку зрения, что как временные, так и пространственные стратегии могут эффективно управлять шумами по формировании паттерна ткани.
Для внеклеточных морфогенов, времена сигнальной интеграции, скорее всего, значительно дольше, чем для Bicoid, это предполагает, что локальные флюктуации в концентрации морфогена могут быть не особенно важными, особенно если сравнивать с другими источниками шумов: рецепторами. Количества рецепторов могут варьировать от клетки к клетке и даже среди клеток с идентичными количествами рецепторов, расположение рецепторов может варьировать из-за стохастической природы связывания и развязывания (Lauffenburger and Linderman, 1993). Как шумы генной экспрессии, так и шумы связывания морфогена с рецепторами д. обнаруживать тенденцию к изменчивости на медленной временной шкале, создаваемые в одном случае с помощью характеристик взрывной транскрипции и трансляции и оборота мРНК и белка, а в др. случае с помощью скорости оборота связанных рецепторов. Поскольку пределы, в которых распространяется градиент, также зависят от скорости потребления и оборота морфогена, то было предположено, что для градиентов морфогенов типичной биологической шкалы длин (50-200 µm ), шумы связывания рецепторов должны быть слишком медленными, чтобы устраняться с помощью простого усреднения во времени (Lander et al., 2009b).
Noise and Tradeoffs
Если эффекты шумов, влияющие на точность формирования паттерна, нелегко удалить с помощью усреднения времени, то как же они преодолеваются? Взгляд на этот вопрос в перспективе forward engineering проливает свет не только на то, как исполнение управляет эволюцией сложной биологической регуляции, но и также на важность выбора оптимальных соотношений-проблема, которая контролирует один аспект работы, часто устраняет другой (Figure 5). Чтобы увидеть это, важно распознать, что неопределенность позиции, создаваемая шумами в интерпретации градиента морфогена, зависит не только от характера шума, то и также от шкалы длин (крутизны) градиента. Это происходит из-за того при крутом градиенте, что расстояние, на котором клетки могут различать, находятся они в одном положении или в другом, более короткое.
Т.о,. сделать градиент более устойчивым к помехам, это сделать его более крутым. Однако из любой данной стартовой амплитуды сделать градиент более крутым, значит укоротить его пределы. В принципе, увеличение стартовой амплитуды градиента морфогена (т.e., продукция больше
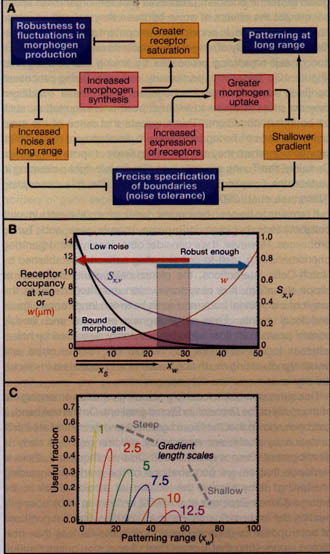 Figure 5. Performance Tradeoffs and Morphogen Gradients
Figure 5. Performance Tradeoffs and Morphogen Gradients
(A) For simple morphogen gradients formed by diffusion with constant receptor-mediated uptake, the ability to achieve performance objectives (e.g., robustness to uncertainty in morphogen production rate; positional precision; and patterning range) is constrained by unintended side effects of performance-enhancing strategies, such as altering levels of morphogen and receptor expression or function.
(B) These tradeoffs may be analyzed quantitatively, by calculating robustness and precision as a function of distance and gradient range. S" is the sensitivity of position to the rate of morphogen production; w is the size of the window of imprecision due to ligand-binding noise. The filled box shows the "useful fraction" of a morphogen gradient, where performance constraints on both Sx," and w are met.
(C) Parameter space exploration suggests that there is some distance beyond which a simple morphogen gradient cannot simultaneously achieve robustness to morphogen synthesis rate, and positional precision, at any location. Useful fractions are plotted as a function of patterning range for various values of gradient length scale. Panels B and C are adapted from Lander et al. (2009b).
морфогена) д. расширить его пределы, но эта стратегия ограничивается др. проблемой: насыщенностью рецепторами. Оказалось, что значительное насыщение рецепторами вблизи источника градиента морфогена драматически снижает устойчивость (robustness) так что незначительные изменения в скорости продукции морфогена (напр., обусловленные средовой и генетической изменчивостью) вызывают крупные изменения в форме градиента (Lander et al., 2009b). Выбор оптимального соотношения между насыщенностью рецепторами на одном конце градиента морфогена и шумами на др. конце является вынужденным с помощью биохимии связывания лиганда и числом рецепторов на клетку. Стратегии преодоления этих ограничений создают дополнительные проблемы. Увеличение количества рецепторов на клетку увеличивает отлавливание морфогена, но укорачивает градиент. Усиление деструкции морфогена может вызывать произвольную устойчивость к флюктуациям в уровнях морфогена (Eldar et al., 2003), но поскольку это делает градиенты мелководными вдали от источника морфогена, то это заканчивается увеличением эффекта шумов на точность (Lander et al., 2009b). То, о чем говорит нам forward engineering, это то, что эксплуатационные качества, связанные с точностью, устойчивостью и размером паттерна, как можно ожидать, всегда будут взаимодействовать др. с др. (напр. Figure 5A). Недавно было предположено (Lander et al., 2009b), что такой выбор оптимального соотношения делает более правдоподобным объяснение для относительно коротких расстояний (50-100 клеток) (Wolpert, 1969), на которых действуют морфогены, чем для физических ограничений скорости распространения морфогенов (Crick, 1970). Учитывая это, мы можем попытаться reverse engineer некоторых из сложных механизмов, наблюдаемых в градиентах морфогенов, чтобы определить может ли какой-нибудь из них помочь с этими компромиссами (tradeoffs). Напр., часто наблюдается, что внеклеточные морфогены накапливаются в везикулярных структурах внутри чувствительных клеток. Если они продолжают передавать сигналы из таких мест-как на самом деле делает это Dpp (Bokel et al., 2006)-то это должно позволять клеткам достигать сигнальной интеграции через определенное время значительно дольше, чем это диктуется скоростью отлавливания морфогенов (Aquino and Endres, 2010). Это объяснение для внутриклеточного накопления морфогена создает альтернативу всё ещё спорной гипотезе, что эндоцитоз играет активную роль в транспорте морфогена.
Др. интересный феномен, способность передачи сигналов Hedgehog (Hh) отражать соотношение связанных к свободным рецепторам скорее, чем количество связанных рецепторов (Casali and Struhl, 2004), это также может служить в качестве стратегии снижения шумов, т.к. это автоматически сокращает эффекты временных флюктуаций в количестве рецепторов.
Spatial Control of Noise
Тот факт, что пространственные характеристики шумов в конечном итоге те, что снижают точность градиента морфогена, указывает на то, что мы д. также искать стратегии снижения шумов, которые точно пространственные. Напр., система градиента Bicoid, тот факт, что ген мишень для Bicoid Hunchback (Hb) сам по себе дает диффундирующую молекулу, которая позволяет эффекты флюктуаций в передаче сигналов Bicoid в одном ядре усреднять для многих ядер (Erdmann et al., 2009; Gregor et al., 2007; Okabe-Oho et al., 2009). Диффузия Hb увеличивает точность путем сглаживания флюктуаций Bicoid, но она также снижает точность путем создания границы Hb менее крутой, компромисс, который ведет к предсказанию максимального эффекта при оптимальной диффузии (Erdmann et al., 2009). Общая стратегия использования градиента морфогена для запуска вторичных процессов, которые, из-за их участия в диффузии, сглаживают пространственные шумы и могут быть обнаружены в системах с внеклеточным морфогеном. Напр., в крыловом диске Drosophila передне-задняя позиционная информация сначала осуществляется с помощью градиента Hh, который действует в коротких пределах, чтобы индуцировать дальнодействующий Dpp градиент (Zecca et al., 1995). Любая неточность в коротких пределах будет интерпретироваться клетками в терминах Hh градиента и будет сглаживать градиент Dpp.
Пространственное усреднение с помощью диффузии не нуждается в использовании для индукции новых морфогенов. Индукция диффундирующих ингибиторов или ко-регуляторов может оказывать сходный эффект. В самом деле, недавнее исследование на крыловых дисках дрозофилы показало, что передача сигналов с помощью градиента Wg может индуцировать продукцию, по крайней мере, двух типов диффундирующих ингибиторов (Piddini and Vincent, 2009).
Особенно мощной стратегией для преодоления пространственных шумов является использование того, что может быть названо "triggered self-organization." В литераутре по биологии развития самоорганизующиеся паттерны обычно призывают в качестве средства для генерации повторяющихся структур- напр., полей из точек или полос. Однако использование дальнодействующего градиента морфогена в качестве импульса для процесса, который закладывает паттерн Turing (напр., экспрессию активатора или ингибитора), оказывается возможным запуск формирования одиночного перехода или пика в специфическом месте пространства (Koch and Meinhardt, 1994). Такой самоорганизующий процесс управляется с помощью диффузии (информации, если не всегда молекул), он должен стремиться к усреднению пространственных шумов на шкале расстояний, связанных с параметрами самого процесса (Figure 4C). Для некоторых самоорганизующихся процессов-таких как паттерны, которые возникают в результате Notch-Delta-обусловленной латеральной ингибиции-точность формирования паттерна может, по-видимому, быть даже улучшена путем добавления пространственных и временных шумов (Cohen et al., 2010).
Образование жилок во время куколочной стадии развития крыльев мух иллюстрирует идею сотрудничества между дально-действующими морфогенами и локальной самоорганизацией. Первоначально градиент Dpp в личиночном крыловом диске устанавливает зачатки крыловых жилок довольно-таки неточно. Позднее, инициация внутри этих зачатков серии событий с использованием коротко-действующей активации и дально-действующей ингибиции или истощения (используя передачу сигналов Notch, Dpp и EGF) (Blair, 2007; Yan et al., 2009) приводит к образованию узких жилок по центрам этих примордиев. Доказательства, что используется процесс Turing, получены при анализе мутаций, которые расширяют зачатки жилок. В этом случае то, что наблюдается, не является более широкими жилками, а дополнительными жилками на той же самой территории (Biehs et al., 1998); это ожидаемый результат, когда домен, поверх которого запускается процесс Turing оказывается крупнее по сравнению с природной длиной волны. Др. ситуация, при которой самоорганизация сужает домен первоначально специфицированный широко с помощью Dpp у эмбрионов Drosophila, где дально-действующий транспорт Dpp в направлении дорсальной стороны эмбриона продуцирует широкий пик передачи сигналов Dpp, который в свою очередь запускает процесс, который усиливает передачу сигналов Dpp в коротких пределах, но ингибирует его в длинных пределах (Umulis et al., 2006; Wang and Ferguson, 2005).
Tradeoffs between Self-Organization and Boundary-Organized Control
Паттерны, продуцируемые с помощью самоорганизации довольно нечувствительны к наружным позиционным сигналам. Эта нечувствительность является обязательной, когда целью формирования паттерна является позиционирование новых событий в отношении мест более ранних событий, особенно, если такое взаимоотношение нуждается в установлении посредством обратной связи. Напр., автоматическое масштабирование процессов Turing трудно достижимо (see, e.g., Ish-ihara and Kaneko, 2006; Othmer and Pate, 1980; Umulis et al., 2008). По этим причинам дально-действующие градиенты морфогена действуют намного лучше. Напротив, процессы, которые самоорганизуются в пространстве противодействуют эффектам шумов, поскольку они естественным образом усредняют пространственную информацию. В принципе развитие может пожинать успехи масштабируемости дально-действующих градиентов и уменьшения шумов самоорганизующихся процессов путем сцепления обоих вместе (Figure 4C). Это работает особенно хорошо с процессами Тьюринга, ограниченных малыми доменами, поскольку в этих условиях они наиболее чувствительны к внешней позиционной информации, такой как пограничные условия. Это указывает на то, что некоторые из наиболее превалирующих, использующих процессы Тьюринга в развитии, могут вовлекать ситуации, в которых они не будут формировать полей из точек и полос. Несомненно, процессы Тьюринга, которые закладывают формирование лево-правостороннего паттерна у позвоночных (создавая одиночную границу), согласуются с этим описанием (Nakamura et al., 2006).
Остается посмотреть, сколь много онтогенетических событий участвуют в совместной работе дальнодействующих морфогенов и процессов Тьюринга. Однако, если мы примем во внимание др. локальные самоорганизующиеся феномены (напр., сигнальные сети между клетками, устанавливаемые с помощью взаимодействий Notch-Delta, путь планарной клеточной полярности ли передачу сигналов Hippo), то легко можем вообразить морфогенез как результат непрерывных передач взад-и-вперед между дальнодействующими сигналами и локальными событиями. Дальнодействующие сигналы оберегают и гибко контролируют позиционную информацию, но легко деградируют с помощью шумов, тогда как локальные события менее изменчивы, но удаляют шумовые и поддерживающие сигналы, действуя подобно усилителю и ретрансляционному устройству (boosters and repeaters) в электрических сетях и при беспроводной трансмиссии.
Эта точка зрения позволяет решить кажущийся парадокс, возникающий в исследованиях градиента Bicoid у дрозофилы. С одной стороны, было показано, что градиент Bicoid и его непосредственная интерпретация с помощью ядер удивительно точна относительно раннего развития (Gregor et al., 2007). С др. стороны, имеются доказательства, что некоторые пертурбации градиента-включая "уплощение" его с помощью изменения местоположения мРНК bicoid в эмбрионе (Ochoa-Espinosa et al., 2009) или "волнений" его из-за неравномерного подогрева эмбриона (Lucchetta et al., 2008)-изменяют положение доменов экспрессии передне-задних генов значительно в меньшей степени, чем предполагалось, указывая тем самым, что в значительной степени точность возникает благодаря самоорганизации. Такая самоорганизация может возникать посредством сложных, перекрестно-регулирующихся сетей генов, которые существуют среди каскадов генов, чья экспрессия первоначально запускается с помощью Bicoid (reviewed by Papatsenko, 2009). Как впервые предположили ещё два десятилетия тому назад (Edgar et al., 1989; Lacalli et al., 1988) и подтвердили недавно (Manu et al., 2009a), gap генов перекрестно-регуляторные взаимодействия создают динамические состояния точек притяжения, характерных свойств самоорганизующихся систем. В самом деле, такая самоорганизация, по-видимому, объясняет устойчивость градиента Bicoid к альтерациям в уровне Bicoid level, в то же самое время продуцирует определенные величины масштабирования относительно размеров эмбриона (Manu et al., 2009b). Остается без ответа вопрос, почему, если первичная роль Bicoid заключается в активации системы, которая обеспечивает значительную часть её устойчивости благодаря самоорганизации, то является ли градиент Bicoid таким точным, каким он является? Reverse engineer всегда должен отвечать на такой вопрос, выдвигая предположение, что существуют дополнительные необходимые рабочие характеристики (performance objectives), которые мы неспособны учесть. Некоторые из них могут быть установлены.
Взгляд на градиенты морфогена как пусковой механизм самоорганизации также хорошо согласуется с недавним исследованием градиентов Hedgehog у беспозвоночных и позвоночных и градиентов ретиноевой кислоты в заднем мозге позвоночных. В таких системах растут доказательства, подтверждающие, что судьбы клеток диктуются в первую очередь историей и продолжительностью воздействия морфогене скорее, чем просто стационарным состоянием количества морфогена (Dessaud et al., 2007, 2010; Maves and Kimmel, 2005). В спинном мозге позвоночных Sonic Hedgehog-индуцированное переключение судеб зависит от перекрестно-регуляторных взаимодействий среди транскрипционных факторов, которые, как и в случае с gap генами у эмбрионов дрозофилы, можно рассматривать как продуцирующие серии состояний attractor (точек притяжения) (Briscoe, 2009; Lek et al., 2010). Такая система может быть описана как система, которая является самоорганизующейся во времени, с градиентом морфогена использующим пространственную склонность к процессу. В крыловом диске дрозофилы градиент Hedgehog также использует временной механизм для продукции множественных границ генной экспрессии (Nahmad and Stathopoulos, 2009). Ясно, что представление о дальнодействующих морфогенах прошло длинный путь с момента ранней модели французского флага (Wolpert, 1969, 2011).
Matching Growth to Pattern
Интуитивно ощущается, что изменения в размере ткани, которые могут быть результатом пищевой или средовой изменчивости, должны приводить к компенсаторным изменениям в выработке позиционных сигналов с помощью морфогенов. Менее очевидно, почему обратное также должно быть верным: что количественные изменения в функции морфогена вызывают заметные изменения в росте ткани. Пока действительно все известные морфогены являются регуляторами роста и большинство способствуют росту. Во многих системах рост зависит от экспрессии в большинстве своем от тех же самых морфогенов, которые устанавливают паттерн. Наиболее изученные примеры получены на крыловом диске Drosophila где и Dpp и Wg являются важными позитивными регуляторами роста (Baena-Lopez et al., 2009; Schwank and Basler, 2010). Исследователи давно пытаются объяснить курьезные наблюдения, что клетки в части крылового диска, формирующие паттерн с помощью Dpp и Wg , пролиферируют в виде более или менее униформного паттерна, тогда как морфогены, которые существенны для управления такой пролиферацией, отчетливо градированы от центра к периферии.
Недавние исследования подтвердили, что ответ на эту загадку связан со способом, с помощью которого морфогены взаимодействуют с Fat/Hippo путем. Среди наиболее интересных эффектов является отсутствие клеточной автономности: активность Yki и последующая пролиферация клеток индуцируются на высоком уровне в клетках крылового диска, которые экспрессируют мишени для сигналов морфогена (от Dpp или Wg) на уровнях существенно более высоких или более низких, чем уровни сигналов в их непосредственных соседях (Rogulja et al., 2008; Zecca and Struhl, 2010). В случае Dpp, было предположено, что этот неавтономный эффект, возникающий из-за градированной передачи сигналов Dpp, ведет к градированной экспрессии Fat лиганда Dachsous и его регулятора Four-jointed, который, в свою очередь, ведет к асимметрии расположения Fat поперек каждой клетки крылового диска, величина которой служит в качестве ингибирующего входящего импульса в путь Hippo (Rogulja et al., 2008).
Одним из следствий этого механизма является то, что клетки в поле морфогена воспринимают сигналы пролиферировать, которые зависят от локального наклона градиента морфогена. Для экспоненциально снижающегося градиента, который является прекрасным приближением градиента Dpp в крыловом диске, наклон, измеренный по отношению к концентрации морфогена в каждой точке, д.б. постоянным, объясняя в принципе, как такой градиент д. управлять пространственно униформной пролиферацией.,
Итея, что наклон градиента морфогена может быть использован для контроля пролиферации, является, фактически, довольно старой и была предположена при изучении феномена интеркалярной регенерации. Она означает тенденцию некоторых эмбриональных или взрослых структур отвечать на хирургические манипуляции, с увеличивающейся избирательностью в тех местах, где клетки, которые ранее были удалены др. от др., оказываются соприкасающимися. Согласно гипотезе рост будет происходить, когда наклон градиента позиционной информации от одной клетки к др., превосходит порог. Это в сущности обратное масштабирование паттерна на размер; это составляет масштабирование размера к паттерну. Интересно, что недавнее исследование показало, что интеркалярная регенерация в крыловом диске использует активацию Yki посредством регуляции пути Hippo (Grusche et al., 2010a; Haider and Johnson, 2011; Sun and Irvine, 2011).
Такие наблюдения подталкивают к спекуляции, что может существовать единый объединяющий принцип-измерение и ответ на наклон градированной позиционной информации-лежащий в основе роли пути Hippo в контроле роста. К сожалению, это почти определенно слишком упрощено. Dpp безусловно влияет на передачу сигналов Yki посредством комбинации клеточно-автономных эффектов (которые зависят от уровня, а не наклона градиента) и неавтономных эффектов (Rogulja et al., 2008; Schwank et al., 2011). И хотя эффекты Dpp's на экспрессию Fat лиганда Dachsous и его регулятора Four-jointed могут объяснить наблюдаемые пролиферативные эффекты, когда клетки принуждаются к контактированию с соседями, которые отличаются существенно своими уровнями Dpp ответа, то представляется, что формы эндогенных градиентов Dachsous и Four-jointed в крыловом диске являются , скорее всего, независимыми от Dpp (Schwank et al., 2011). Сходным образом, в то время как ранние исследования указывали на то, что пролиферативные ответы на различия между соседними клетками в передаче сигналов Wg (который индуцирует активность Yki) (Zecca and Struhl, 2010) может быть основным средством, с помощью которого градиент Wg управляет ростом (Baena-Lopez and Gar-cia-Bellido, 2006), последняя работа показала, что униформные, умеренные уровни Wg являются мощными стимулами для пролиферации и далее, что наклон градиента Wg является слишком незначительным в большей части крылового диска, чтобы вызывать неавтономные эффекты (Baena-Lo-pez et al., 2009). Дополняет эти наблюдения недавняя работа Schwank et al. (2011), показывающая, что передача сигналов Fat/Hippo может быть градированной вдоль передне-задней оси крылового диска, даже когда отсутствует явный градиент морфогена с этом направлении. Хотя Schwank et al. полагают, что передачи сигналов Fat/Hippo и Dpp являются независимыми от путей контролирующих рост, которые действуют в разных доменах диска, другим объяснением является то, что передача сигналов Hippo интегрирует позиционную информацию, которая исходит из источника, ещё неучтенного для любой современной модели.
Чтоб источник мог быть механической силой. Как описано ранее, существуют строгие указания, что натяжение и сдавливание внутри эпителия крылового диска играют роль в контроле роста (Aegerter-Wilmsen et al., 2007, 2010; Hufnagel et al., 2007). Недавние исследования двойного лучепреломления при деформации (stress-birefringence) показали присутствие градиента сдавливания от центра к периферии в крыловых дисках (Nienhaus et al., 2009). Учитывая тесную связь между вышестоящими компонентами пути Hippo и компонентами клеточных соединений и цитоскелета, было предположено, что путь Hippo непосредственно воспринимает механические импульсы (Grusche et al., 2010b). Очевидно, что имеется настоятельная необходимость в экспериментальном выяснении роли механических событий на путь передачи сигналов Hippo.
C др. стороны, этот источник может исходить из самих градиентов морфогена, посредством механизмов, родственных их способности запускать процессы самоорганизации в специфических местах. Как уже подчеркивалось, Wg ген мишень vestigial (vg), который, по-видимому, играет роль в управлении ростом крылового диска, способен к аутоактивации (Zecca and Struhl, 2007, 2010), ещё vg также, по-видимому, является мишенью для коротко-действующей негативной обратной связи посредством неизвестного механизма (Piddini and Vincent, 2009). Аргументы, базирующиеся на математическом моделировании, постулируют, что подобное упорядочение создает ситуацию, при которой установившийся режим баланса между Vg-способствующему росту, Vg-способствующему экспрессии vg и Wg-зависимому подавлению экспрессии vg достигается только при фиксированном размере ткани (Zhu, 2011). Поскольку эта недавняя работа исследовала только ограниченное число избранных параметров, необходимо проверить, оперирует ли этот механизм действительно в крыловых дисках. Тем не менее, он предлагает элегантное решение проблемы достижения set-point контроля роста на относительно длинной шкале расстояний.
Необходимо отметить, что одновременное существование механизмов масштабирования паттерна роста (scale pattern to growth) и роста до паттерна (growth to pattern) д. создавать-если такие механизмы действительно независимы один от др. -бесполезные циклы, при этом каждый из процессов постоянно управляет другим. Ясно, что подобные механизмы не могут быть независимыми, но как они связаны, остается неясным. Одной из интригующих возможностей является то, что используются glypicans клеточной поверхности, такие как Dally и Dally-like, которые, как было установлено, являются прямыми мишенями сигнального пути Hippo (Rodriguez et al., 2008). Эти молекулы и из ортологи у млекопитающих строго участвуют как в контроле размера органа (Filmus and Capurro, 2008; Selleck, 1999; Takeo et al., 2005) , так и в регуляции формирования паттерна морфогенетических градиентов (e.g., Belenkaya et al., 2004; Franch-Marro et al., 2005; Galli et al., 2003; Han et al., 2004, 2005; Kreuger et al., 2004).
Lessons and Implications
The remarkable regulation, canalization, robustness, and precision of embryonic development suggest that developing systems devote a considerable amount of cellular machinery to the explicit purpose of control. Through forward and reverse engineering, it has become possible to systematically explore some of the control challenges faced by developing embryos and link such challenges to enabling mechanisms. In true systems biology fashion, such work strives to explain the complexity of developmental mechanisms in terms of the coordinated functions of entire systems, and not just that of individual parts. The studies highlighted above provide general lessons for future research into control of morphogenesis and development. Three of the most salient lessons are described below. (1) Performance Is Always Subject to Tradeoffs The engineering dictum that "there's no free lunch" makes the point that control always comes at a cost. Making a system robust in one way invariably makes it fragile in another (Doyle and Csete, 2007). Tightly controlling cell number can hamper regeneration speed; controlling how robust morphogen gradients are to fluctuations in morphogen levels can affect how sensitive they are to noise; using spatially self-organizing processes to suppress the effects of noise can make spatial scaling more challenging. In general, the researcher who proposes that a particular mechanism fulfills a particular control function should consider how other types of performance are degraded as a result of such control.
Unfortunately, this is a standard not often met in the current literature, even in journals with a strong systems biology focus.
(2) Control Is Not Micromanagement
It is easy to think that, in order for a system to be under tight control, every part of it must be controlled. Yet the strategy of integral feedback (Figure 3) shows that extremely tight control can be achieved merely by feeding back the right kind of signal at one point in a network into an earlier point. The intervening dynamics needn't be subject to any special regulation (even though it may appear that they are). For example, as long as there is any sort of feedback control on the renewal probabilities of stem cells, such cells can be wildly stochastic in their individual behaviors (e.g., Chang et al., 2008; Clayton et al., 2007; Gomes et al., 2011; Snippert et al., 2010) yet still give us the impression that they "know" precisely what they are doing. Failure to recognize that tightly controlled systems can include uncontrolled parts may well explain the discomfort that many biologists have long had with random processes, such as stochastic cell-fate switches (Chang et al., 2008), and the idea that mere diffusion creates morphogen gradients. Recent assertions that diffusion is "too messy" (i.e., too hard to control) to get the job done (Wol-pert, 2009,2011) indicate that confusion between micromanagement and control is very much alive in biology. In fact, because control always comes at a price, systems that achieve it without micromanaging are often better off.
(3) Phenotype Is Not Performance
Experimental genetics has become an indispensible tool of developmental biology because it enables us to infer causal connections between mechanisms (gene activity) and observations (phenotypes). Impressed with the intricate beauty of the networks that such approaches construct, it is easy to lose sight of the fact that what evolution selects for is not phenotype per se but performance, the ability of phenotype to do something useful. Too often, the phenomena we choose to investigate are selected for study based on criteria that may be only obliquely related to actual importance to the organism. For example, the high precision of the Drosophila Bicoid gradient is fascinating to us, but we still don't know how much it matters to the fly. In the wing disc, numerous studies have focused on explaining how spatially graded morphogens drive spatially uniform growth, when we actually have no evidence that the uniformity of growth is itself particularly important. From a traditional molecular biology standpoint, these are not serious problems because investigating these phenomena will likely still lead us to new mechanisms. But for the systems biologist, who seeks to use notions of design and performance to place mechanisms into context, there is a real need to get a systematic handle on what is phenomenon and what is epiphenomenon. Luckily, this is just what the tools of forward and reverse engineering provide. As our knowledge of the mechanistic complexity of developmental regulation grows, we can expect to see such approaches playing an ever-greater role in making sense of it all.
|
Сайт создан в системе
uCoz)