Миллиарды нейронов образуют точные соединения по всей нервной системе позвоночных. Чтобы установить такие соединения аксоны нервных клеток д. находить свои пути к соотв. им мишеням во время нервного развития. Пока избирательные пути, которые используются для осуществления такой проводки в головном мозге, изучены недостаточно. Сигналы. которые регулируют наведение аксонов многочисленны и изменчивы. Сюда входят те, что диффузны или закреплены, те, которые действуют на далеких и близких расстояниях и те, которые отталкивают, притягивают и/или адгезивны по отношении к растущим аксонам (Yamamoto et al., 2003, Chilton, 2006; Erskine and Herrera, 2007). Помимо идентификации сигналов ведения аксонов и соотв. им рецепторов, существуют внутриклеточные механизмы, которые участвуют в росте, нахождении пути и мишеней аксонами. Сигналы наведения, как полагают, контролируют поведение аксонов посредством посредством переустройства цитоскелета, это может управлять динамикой у наведением ростового конуса (Dent and Ger-tler, 2003; Farrar and Spencer, 2008; Lowery and Van Vactor, 2009; Bashaw and Klein, 2010).
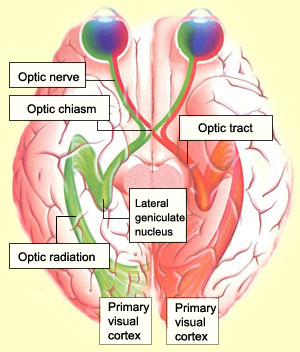
Retinotectal путь у Xenopus laevis служит в качестве важной модели ведения аксонов. Значительная часть нейрогенеза и нейроанатомии Xenopus retinal ganglion cells (RGCs) хорошо известна и эти клетки создают единственную связь между глазом и головным мозгом, которая обеспечивает зрительное восприятие (Holt, 1989; Harris and Holt, 1990; Dingwell et al., 2000). Стереотипический путь, который RGC аксоны проходят от сетчатки к среднему мозгу, хорошо охарактеризован. Xenopus, в частности, прекрасная модель из-за доступности RGC аксонов по всей длине зрительного пути и из-за относительной легкости, с которой могут быть получены эмбрионы Xenopus и подвергаться манипуляциям (Chien and Harris, 1994). Ретинальные аксоны проходят через совершенно различные микроусловия во время развития зрительной системы. Они сначала простираются внутри сетчатки к оптическому диску и головке нерва, образуя пучок аксонов, зрительный нерв, как только они покидают глаз, начиная со ст. 28 у Xenopus. На ст. 32, эти аксоны подготавливаются, чтобы иннервировать контрлатеральную сторону головного мозга путем пересечения эмбриональной срединной линии, образуя тем самым оптический перекрест во время прокладки контрлатеральных путей. Ретинальные аксоны затем идут вдоль боковой поверхности нейроэпителия диэнцефалона, образуя зрительный тракт, прежде чем повернуть в каудальную сторону (на ст. 35/36), в направлении своей финальной мишени в конце тракта, зрительного tectum, на ст. 39/40. Вдоль всего этого пути ростовые конусы RGC д. производить оценку своего окружения, чтобы найти соотв. молекулярные сигналы, которые будут наводить их движения и приводить их к соотв. финальными мишеням для последующего синаптогенеза в зрительном tectum.
Недавние доказательства подтвердили, что классические морфогены, такие как члены семейств Wnts, TGFβ/BMP и Hedgehogs, могут служить в качестве сигналов ведения различных аксонов у разных организмов (Charron and Tess-ier-Lavigne, 2005; Sanchez-Camacho et al., 2005; Zou and Lyuksyutova, 2007). Несколько групп предоставили доказательства, что Sonic hedgehog (Shh) является важным сигналом ведения аксонов в ходе развития позвоночных. У эмбрионов кур Shh важен для нахождения пути внутри сетчатки (Kolpak et al., 2005) и к оптическому перекресту (Trousse et al., 2001). У мышей Shh действует во время расширения RGC внутри сетчатки (Sanchez-Camacho and Bovolenta, 2008), и в наведении на оптический перекрест (Hao et al., 2007; Sanchez-Camacho and Bovolenta, 2008). Shh также используется комиссуральными аксонами в спинном мозге мышей, чтобы содействовать прохождению аксонами срединной линии в сотрудничестве с Netrin-1 (Charron et al., 2003) и с помощью Boc рецепторов (Okada et al., 2006). Пост-комиссуральное ведение аксонов вдоль продольной оси спинного мозга также использует Shh сигналы (Bourikas et al., 2005). В комиссуральных аксонах крыс неканоническая передача сигналов Shh посредством киназ семейства Src была описана в качестве механизма для аксонов, чтобы избегать транскрипции и это позволяет быстро отвечать на сигналы наведения Shh в конусах роста (Yam et al., 2009).
В целом растут доказательства, что Shh является важным сигналом ведения различного типа аксонов. В зрительной системе, как показано выше, Shh описывается как хемоаттрактант и репелент в сетчатке и как репелент в ретинальных аксонах в оптическом перекресте. После этих точек вдоль зрительного пути ретинальные аксоны могут всё ещё перемещаться на относительно длинные расстояния (approximately 430 µm у Xenopus; Harris et al., 1985) чтобы достигнуть своей финальной мишени, но далее сигналы наведения вдоль зрительного тракта не достаточно охарактеризованы. Предыдущие исследования показали, что ретинальные аксоны Xenopus базируются на направляющих сигналах, которые являются стабильными и локальными для зрительного тракта (Harris, 1989; Taylor, 1990; Cornel and Holt, 1992), а молекулы, такие как heparan sulfates (Walz et al., 1997; Irie et al., 2002) и матричные metalloproteinases (Webber et al., 2002; Hehr et al., 2005: Chen et al., 2007) принимают участие в этом процессе. Роль морфогенов, таких как Shh в диэнцефалоне оставалась неизвестной. Поэтому было решено проверить, участвует ли передача сигналов Hedgehog в ведении ретинальных аксонов в регионах после пересечения ими срединной линии, когда RGC проецируют свои аксоны из вентрального в дорсальные концы развивающегося диэнцефалона. Была выявлена новая функция Sonic hedgehog в качестве сигнала наведения для ретинальных аксонов вдоль зрительного тракта у Xenopus. Было также показано, что передача сигналов Hedgehog участвует в распознавании мишеней зрительного tectum.
Сайт создан в системе
uCoz 