|
|
|
|---|---|
Loss of Lhx1 Activity Impacts on the Localization
of Primordial Germ Cells in the Mouse
 Developmental Dynamics V.23: P.2851-2859, 2010 |
|
|
Mouse embryos lacking Lhx1 (Lim1) activity
display defective gastrulation and are deficient of primordial germ
cells (PGCs) (Tsang et al. [2001] International Journal of
Developmental Biology 45:549-555). To dissect the specific role of Lhx1
in germ cell development, we studied embryos with conditional
inactivation of Lhx1 activity in epiblast derivatives, which, in
contrast to completely null embryos, develop normally through
gastrulation before manifesting a head truncation phenotype. Initially,
PGCs are localized properly to the definitive endodenn of the posterior
gut in the conditional mutant embryos, but they depart from the
embryonic gut prematurely. The early exit of PGCs from the gut is
accompanied by the failure to maintain a strong expression of Ifitm1 in
the mesoderm enveloping the gut, which may mediate the repulsive
activity that facilitates the retention of PGCs in the hindgut during
early organogenesis. Lhx1 therefore may influence the localization of
PGCs by modulating Ifitm1-mediated repulsive activity. Developmental
Dynamics Vol.23: P.2851-2859, 2010 Wiley-Liss, Inc.
|
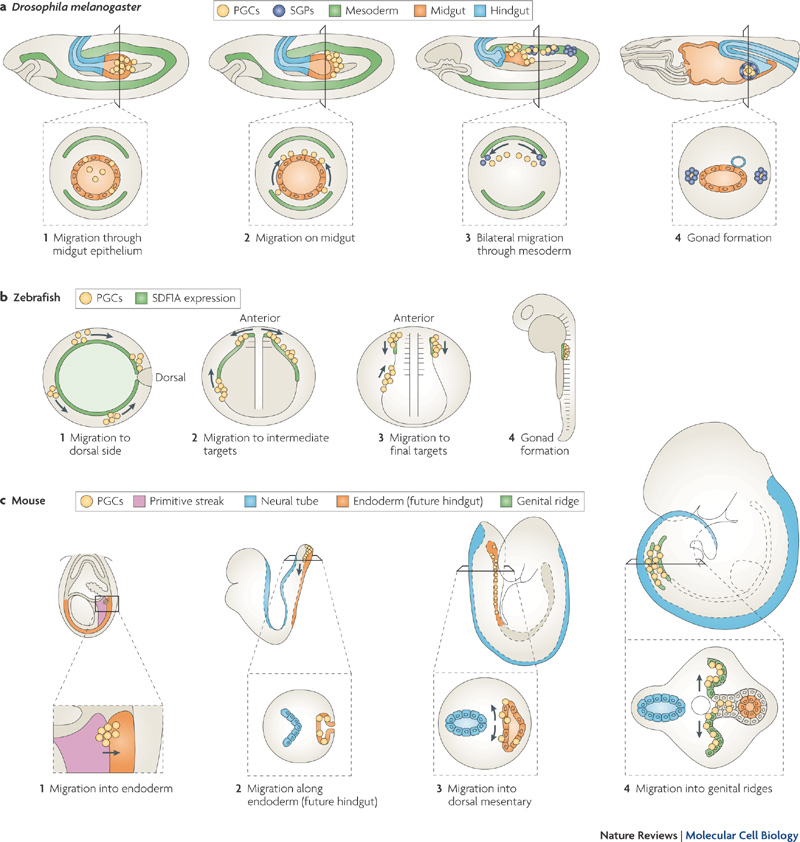 Стадии миграции примордиальных зародышевых клеток У мышей информация о развитии зародышевых клеток получена в
результате анализа влияния поврежденной активности генов ан
спецификацию, пролиферацию и миграцию primordial germ cells (PGCs).
Напр., эмбрионы, которые лишены активности bone mophogenetic protein
(Bmp)-4 , не активируют Prdml (Blimpl) в эпибласте (Ohinata et al.,
2005) и полностью лишены PGCs (Lawson et al., 1999). После образования
PGCs локализация PGCs в разных зародышевых слоях зависит от экспрессии
interferon induced transmembrane protein (Ifitm)-1 (mil-2/fragilis2) и
Ifitm3 (mil-l/fragilis). Ifitm1, который выявляет гомотипические
межклеточные взаимодействия, экспрессируется в мезодерме у эмбрионов
стадии гаструлы. Предшественники PGC в мезодерме экспрессируют Ifitm1,
но те, что перемещены в энтодерму прекращают экспрессировать этот ген
(Tanaka and Matsui, 2002). Это подавление Ifitm1, как полагают,
генерирует отталкивающее взаимодействие между мезодермой и
предшественниками PGC и облегчает их переход из Ifitm1-экспрессирующей
мезодермы в энтодерму, которая не экспрессирует Ifitm1 (Tanaka et al.,
2005). PGCs , происходящие из этих предшественников, которые
располагаются в энтодерме задней кишки, остаются чувствительными к
Ifitm1-обусловленно отталкивающей активности (Tanaka et al., 2005).
Удержание PGCs в энтодерме кишки до повторной инициации миграции из
кишки через мезентерий в генитальные гребни, может быть ответственно за
реакцию PGCs на репульсивную активность, оказываемую
Ifitm1-экспрессирующей мезенхимой, соседствующей с энтодермой (Tam et
al., 2006).
Стадии миграции примордиальных зародышевых клеток У мышей информация о развитии зародышевых клеток получена в
результате анализа влияния поврежденной активности генов ан
спецификацию, пролиферацию и миграцию primordial germ cells (PGCs).
Напр., эмбрионы, которые лишены активности bone mophogenetic protein
(Bmp)-4 , не активируют Prdml (Blimpl) в эпибласте (Ohinata et al.,
2005) и полностью лишены PGCs (Lawson et al., 1999). После образования
PGCs локализация PGCs в разных зародышевых слоях зависит от экспрессии
interferon induced transmembrane protein (Ifitm)-1 (mil-2/fragilis2) и
Ifitm3 (mil-l/fragilis). Ifitm1, который выявляет гомотипические
межклеточные взаимодействия, экспрессируется в мезодерме у эмбрионов
стадии гаструлы. Предшественники PGC в мезодерме экспрессируют Ifitm1,
но те, что перемещены в энтодерму прекращают экспрессировать этот ген
(Tanaka and Matsui, 2002). Это подавление Ifitm1, как полагают,
генерирует отталкивающее взаимодействие между мезодермой и
предшественниками PGC и облегчает их переход из Ifitm1-экспрессирующей
мезодермы в энтодерму, которая не экспрессирует Ifitm1 (Tanaka et al.,
2005). PGCs , происходящие из этих предшественников, которые
располагаются в энтодерме задней кишки, остаются чувствительными к
Ifitm1-обусловленно отталкивающей активности (Tanaka et al., 2005).
Удержание PGCs в энтодерме кишки до повторной инициации миграции из
кишки через мезентерий в генитальные гребни, может быть ответственно за
реакцию PGCs на репульсивную активность, оказываемую
Ifitm1-экспрессирующей мезенхимой, соседствующей с энтодермой (Tam et
al., 2006). Активности нескольких генов транскрипционных факторов,
таких как Brachyury (T), Foxa2 и Lhx1 (Lim1) необходимы для образования
PGC, хотя молекулярный путь, посредством которого эти факторы могут
действовать, неизвестен (Tsang et al., 2001). Потеря Lhx1 ведет к
отсутствию PGCs у некоторых мутантных эмбрионов, тогда как у др. PGCs
присутствуют в уменьшенных количествах. У некоторых Lhx1-нулевых
эмбрионов кластер позитивных по щелочной phosphatase PGCs
обнаруживается расположенным эктопически во внеэмбриональной мезодерме
вдалеке от первичной полоски (Tsang et al., 2001). Эти клетки вряд ли
вносят вклад в популяцию PGC внутри эмбриона и вряд ли могут
объяснить уменьшение популяции PGC у Lhx1-нулевых эмбрионов (Tsang et
al., 2001). Эти находки указывают на то, что Lhx1 может иметь разные
роли в образовании PGCs и собственно в их локализации. В согласии с
этой последней ролью активность Lhx1, как было установлено, необходима
для морфогенетического перемещения аксиальной мезодермы у эмбрионов Xenopus
и мезодермальных и энтодермальных клеток у эмбрионов мыши (Hukriede et
al., 2003; Shimono and Behringer, 2003; Tam et al., 2004). Смущающим
фактором в анализе влияния потери Lhx1 на образование PGC является то,
что образование зародышевых слоёв также тяжело нарушается у
Lhx1-нулевых мутантных эмбрионов (Tsang et al., 2000). Химерные
эмбрионы с дикого типа внеэмбриональными тканями и Lhx1-нулевым
мутантными производными эпибласта не обнаруживают различимых дефектов в
образовании и морфогенезе зародышевых слоёв, указывая тем самым, что
активность Lhx1 в не-эпибластных производных, таких как висцеральная
энтодерма, необходима для осуществления гаструляции (Shawlot et al.,
1999). В данном исследовании мы выявили специфический эффект потери
функции Lhx1 на PGCs путем преодоления затемняющего картину эффекта
аномальной гаструляции, чтобы исследовать эмбрионы, лишенные функции
Lhx1 преимущественно в эпибласте, посредством Meox2-Cre-обеспечиваемой
эксцизии floxed Lhx1 аллеля (Kwan and Behringer, 2002). Было
установлено, что активность Lhx1 в эпибласте и его производных не
влияет на образование PGCs, но влияет на их пребывание в энтодерме
задней кишки, что впоследствии может приводить к потере PGCs. Мы
полагаем, что преждевременный выход PGCs из задней кишки может быть
связан с неспособностью выявлять необходимую Iftiml-обусловленную
репульсивную активность, которая ограничивает эмиграцию PGCs в
мезодерму, окружающую энтодерму кишки.
|