Perigemmal клетки вкусовых почек, но не сами вкусовые клетки (но см. Asano-Miyoshi et al., 2008), экспрессируют cytokeratins 5 и 14 (K5 and K14) (Lee et al., 2006; Asano-Miyoshi et al., 2008; Okubo et al., 2009), которые также экспрессируются популяцией регенеративных клеток кожи, базальными кератиноцитами общего эпидермиса (Bragulla and Homberger, 2009). Недавно Okubo с коллегами использовали ткане-специфифицированные, индуцибельные молекулярные генетические инструменты для картирования потомства базальных кератиноцитов, экспрессирующих K14, и расположенных во вкусовом эпителии (Okubo et al., 2009). Экспрессия неустраняемого lacZ репортерного гена индуцировалась с помощью tamoxifen у мышей, несущих: 1) один аллель с человеческим K14 промотором (hK14), управляющим слитым белком из Cre recombinase и tamoxifen-чувствительной формой эстрогенового рецептора (Vasioukhin et al., 1999); и 2) второй аллель, в котором конституитивный промотор R26 локуса, управляющий lacZ, находился ниже flox-stop-flox кассеты (Soriano, 1999). Результаты этих исследований мучительные, но неубедительные. Безусловно, hK14-экспрессирующие базальные кератиноциты давали клетки внутри вкусовых почек, также как и папиллярного эпителия и общего эпителия языка. В некоторых случаях потенциально клональные кластеры меченных клеток представляли собой как вкусовую почку, так и клетки сосочка, указывая, что K14+ базальные клетки являются мультипотентными. Однако чтобы решить этот вопрос, необходимо проверить инициально меченную(е) клетку(и) и отследить их производные более тщательно, вообще-то с помощью титрации понижающейся дозы tamoxifen, чтобы метить только очень немногие клетки на животное. Базальные кератиноциты эпителия языка и вкусовых сосочков также экспрессируют транскрипционные факторы Trp63 на высоких уровнях и Sox2 на низких уровнях (Okubo et al., 2009). Экспрессионные характеристики K14
клеток, как было предположено, определяют пул предшественников вкусовой почки. В попытке исследовать потенциал дифференцировки этой популяции клеток, данная группа получила и культивировала популяцию сильно флюоресцентных клеток из эпителия языка мышей, несущих аллель, где промотор человеческого cytokeratin 5 управлял GFP (Luo et al., 2009). Этот подход давал эпителиальные стволовые клетки, которые могли давать дифференцированный эпителий, но, по-видимому, не продуцировали вкусовые почки и располагающиеся в них руцепторные клетки.
Дополнительные новые данные по генезу вкусовых почек получены в исследованиях эмбрионального развития вкусового эпителия. В то время как дифференцировка вкусовых клеток происходит примерно при рождении у крыс и мышей, то первые указания на то, что происходит спецификация этих структур приходятся на средину беременности. У мышей язык формируется из билатеральных парных языковых выростов, которые сливаются по средней линии на эмбриональный день (E) 11.0-11.25. На ст. E12.5, серии фокальных эпителиальных утолщений или плакод, формируются в два билатеральных ряда, предваряющих локализацию грибовидных сосочков взрослых (Fig. 2). Важно, что эти плакодные клетки экспрессируют как мРНК, так и белок Sonic Hedgehog (Shh) на этой ранней ст. развития. Поскольку вкусовые плакоды, как было предположено, представляют собой предшественники сосочков (Mistretta and Liu, 2006), то судьба этих Shh-экспрессирующих клеток не была детерминирована. Применив сходную индуцибельную молекулярную генетическую схему Okubo и коллег, мы использовали мышиные эмбрионы, обладающие аллелем, где нативная Shh кодирующая область была замещена Cre-ERtam слитым белком (Harfe et al., 2004) и Rosa LacZ репортерным аллелем, для картирования судьбы Shh-экспрессирующих вкусовых плакод. Обработав беременных самок tamoxifen точно, когда Shh фокально экспрессируется во вкусовых плакодах эмбриона, мы установили, что эти клетки были предшественниками дифференцированных вкусовых клеток у постнатальных животных, но не вносят вклада в эпителий вкусовых сосочков (Thirumangalathu and Barlow, 2009; Thirumangalathu et al., 2009). Более того, было также показано, что, по крайней мере, type I и II вкусовые почки происходят непосредственно от Shh-экспрессирующих клеток вкусовых плакод, постоянно маркируемых между E12.5 и E14.5. Дополнительные гипотезы, решаемые с помощью нашего исследования , заключались в том, что эмбриональные Shh-экспресирующие вкусовые предшественники д. персистировать у взрослых мышей в качестве стволовых клеток вкусовых почек. Однако это оказалось не так; скорее клетки вкусовых почек, происходящие из Shh-экспрессирующих плакод, постепенно теряются постнатально и полностью отсутствуют спустя 4 мес. после рождения. Учитывая эти находки, мы предположили, что Shh-экспрессирующие плакодные клетки функционируют в качестве сигнального центра у эмбрионов и молодых постнатальных животных и что дополнительные клетки предшественники рекрутируются у взрослых мышей, чтобы перенять необходимую функцию непрерывного пополнения клеток зрелых вкусовых почек, подобный сценарий продемонстрирован для волосяных фолликулов мышей (Nowak et al., 2008). Рекрутирование дополнительных клеток предшественников, экспрессирующих Shh, как полагают, продолжается в базальной области вкусовых почек в течение всей жизни, как было описано далее. Альтернативно, Shh-экспрессирующие стволовые клетки вкусовых почек могут быть заложены раньше или позднее в эмбриогенезе, так что исследования по картированию судеб, осуществляемые в более ранние или более поздние моменты времени, будут выявлять эту стволовую популяцию. Ведутся исследования для тестирования этих разных идей. Shh экспрессируется также во взрослых вкусовых почках мышей. Помимо веретенообразных вкусовых клеток types I, II и III, идентифицирован 4-й тип клеток; это округлые клетки в базальном компартменте вкусовых (Murray, 1973; Royer and Kinnamon, 1991; Taniguchi et al., 2005), в большинстве, если не во всех типах IV, клетках, как полагают, экспрессируется Shh (Miura et al., 2001; Miura et al., 2003; Miura et al., 2004; Miura
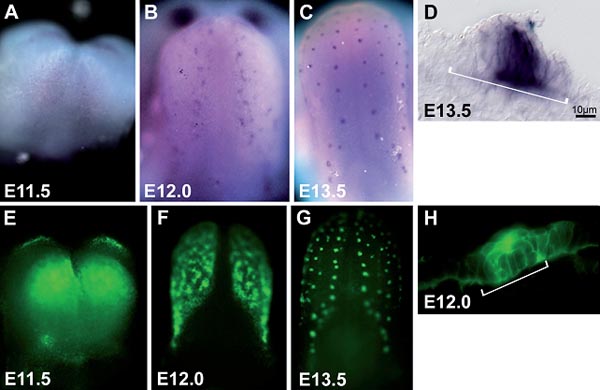 Fig. 2. - Shh expression in developing tongues. In situ hybridization (A-D) and protein expression (E-H) reveal Shh throughout the lingual epithelium at E11.5 (A, E). Shh resolves at E12.0 (B, F, H), and is restricted to the taste placodes by E13.0 (C, D, G). A-C. Shh mRNA (purple) in whole tongues at progressive developmental stages. D. An E13.5 tongue cryosection reveals Shh in a subset of epithelial cells within a developing taste papilla (white bracket). E-G. Whole mount tongues immunofluorescently stained for Shh protein (green) exhibit the same expression pattern as Shh mRNA. H. An immunostained tongue cryosection at E12.0 reveals Shh expression is coincident with the morphological extent of the placode (white bracket). Anterior is up A-C, E-G. Scale: D and H = 10 µm. Images courtesy of Dr. Shoba Thirumangalathu.
Fig. 2. - Shh expression in developing tongues. In situ hybridization (A-D) and protein expression (E-H) reveal Shh throughout the lingual epithelium at E11.5 (A, E). Shh resolves at E12.0 (B, F, H), and is restricted to the taste placodes by E13.0 (C, D, G). A-C. Shh mRNA (purple) in whole tongues at progressive developmental stages. D. An E13.5 tongue cryosection reveals Shh in a subset of epithelial cells within a developing taste papilla (white bracket). E-G. Whole mount tongues immunofluorescently stained for Shh protein (green) exhibit the same expression pattern as Shh mRNA. H. An immunostained tongue cryosection at E12.0 reveals Shh expression is coincident with the morphological extent of the placode (white bracket). Anterior is up A-C, E-G. Scale: D and H = 10 µm. Images courtesy of Dr. Shoba Thirumangalathu.
et al., 2005b). Эти клетки являются предположительно популяцией преходящих клеток предшественников вкусовых почек, которые впервые появились в эмбриональных Shh-экспрессирующих вкусовых плакодах (Nakayama et al., 2008). Shh-экспрессирующие type IV клетки не являются стволовыми клетками вкусовых почек, т.к. они подобно веретенообразным вкусовым клеткам являются постмитотическими (Miura et al., 2004). Shh-экспрессирующие базальные клетки могут представлять собой раннюю фазу дифференцировки незрелых вкусовых почек, учитывая, что BrdU пульсовое мечение Shh-экспрессирующих intragemmal базальных клеток достигает пика приблизительно 24 ч после их последнего perigemmal деления (Miura et al., 2004). Поскольку качественные особенности вкусовых клеток, производных Shh-экспрессирующих type IV клеток, не были определены с полной определенностью, то наши предварительные данные указывают на то, что, по крайней мере, type II клетки продуцируются этим клоном (H. Miura, J. Liang and L.A. Barlow, unpublished results).
Помимо своей экспрессии на дискретных стадиях дифференцировки вкусовых клеток Shh может также функционировать внутри вкусовых почек как регулятор обновления клеток. Напр., Shh регулирует рост волос и формирование перьев, а также участвует в гомеостазе кожи (Sato et al., 1999; Chuong et al., 2000; Stenn and Paus, 2001; McKinnell et al., 2004; Athar et al., 2006). В этих тканях Shh действует как позитивный регулятор пролиферации клеток и может выполнять сходную роль во вкусовом эпителии, в особенности, поскольку Ptch1-экспрессирующие чувствительные к Shh клетки располагаются в perigemmal пролиферативной зоне, окружающей каждую почку (Miura et al., 2001). Базальные клетки вкусовых почек, по-видимому, обладают некоторыми общими молекулярными и клеточными свойствами с клетками вкусовых плакод: и те и др. экспрессируют не только Shh, но и Prox1 (Nakayama et al., 2008); , по-видимому, они являются промежуточными клетками предшественниками вкусовых почек; и митотически молчащими (Mbiene and Roberts, 2003). Однако заслуживают внимания чёткие различия между ними: 1) экспрессия Shh в базальных клетках строго зависит от вкусовых нервов у взрослых, тогда как Shh-экспрессирующие вкусовые плакоды появляются до иннервации; 2) Cytokeratin 8(CK8)-подобная иммунореактивность присутствует в веретенообразных, но не в базальных клетках во вкусовых почках (Knapp et al., 1995), тогда как вкусовые плакоды являются CK8 позитивными (Mbiene and Roberts, 2003); и 3) временное окно экспрессии
Shh во вкусовых плакодах, по-видимому, более продолжительное, чем в базальных клетках вкусовых почек. Неясно когда и как появляются эти различия в Shh-экспрессирующих клетках предшественниках вкусовых почек во время развития. Некоторые намеки, впрочем, исходят от периода чувствительности зависимости от нервов вкусовых почек (Hosley et al., 1987). Количество вкусовых почек, развивающихся в circumvallate сосочках крыс, существенно зависит от иннервации в течение 10 дней после рождения, указывая тем самым, что вкусовые нервы могут участвовать в превращении Shh-экспрессирующих клеток из эмбрионального во взрослое состояние. Ряд др. сигнальных факторов и регуляторныех генов экспрессируется во взрослых вкусовых почках, но их функция, кстати, не установлена. Предполагается, что продукты этих генов могут использоваться, исходя из их функции в др. эпителиальных и нервных системах. Помимо Shh, путь Notch (N) участвует в обновлении вкусовых клеток, исходя из паттернов экспрессии N receptors 1, 2 и 4, некоторых трансмембранных N лигандов, включая Delta-like 1 (Dll-1) и Jagged 1 и 2, и известные транскрипционные факторы этой сигнальной системы, т.e., Mash1, Hes6 и Hes1 (Seta et al., 2003; Miura et al., 2005a; Seta et al., 2006a; Seta et al., 2006b; Ota et al., 2009). Экспрессия
Mash1 чётко ассоциирует с клетками типа III cells (Seta et al., 2006b) и может также экспрессироваться субнабором клеток, которые в конечном итоге дифференцируются во вкусовые клетки типа II (Miura et al., 2005a). Т.о., путь Notch может регулировать выбор судеб клетками во время обновления вкусовых клеток. Sox2 является транскрипционным фактором, важным для эмбрионального развития и может действовать как "master regulator" мультипотентности стволовых клеток в ряде тканей (Rizzino, 2009). Во вкусовой системе Sox2 необходим для развития эмбриональных вкусовых почек (Okubo et al., 2006) и экспрессируется также во вкусовых почках взрослых, как в незрелых вкусовых почках, так и дифференцированных клетках типа I (Suzuki, 2008). Sox2 экспрессируется также в нервных предшественниках развивающейся и взрослой нервной системы (Ellis et al., 2004), где он участвует в поддержании состояния нервных предшественников и регулирует судьбу нервных клеток (Bylund et al., 2003; Graham et al., 2003; Episkopou, 2005). Принимая во внимание, что Sox2 экспрессируется как intragemmally, так и perigemmally во вкусовых органах взрослых (Okubo et al., 2006; Suzuki, 2008; Okubo et al., 2009), этот транскрипционный фактор обладает рядом функций в обновлении вкусовых клеток.
Taste bud homeostasis and the response(s) to injury
В нормальных условиях регенерация вкусовых клеток скорее всего происходит по соотв. этапам. Однако эти сенсорные клетки чувствительны к повреждениям и вкусовые почки сами по себе испытывают влияния со стороны ряда сценариев, которые имеют отношение к здоровью человека. Вообще-то, большинством тщательно исследованных образцов при исследовании морфологии вкусовых почек является модель денервации. Известно, что дифференцировка вкусовых почек зависит от интактной иннервации (Vintschgau and Honigschmied, 1876; Torrey, 1934; Guth, 1958; Sollars et al., 2002; Oakley and Witt, 2004); когда или веточка chorda tympani нерва VII или glossopharyngeal нерв (краниальный ганглий IX) перерезается, то грибовидные вкусовые почки передней части языка или вкусовые почки circumvallate сосочков, соотв., редуцируются. Влияние денервации на вкусовые почки грибовидных сосочков, по-видимому, менее сильное, чем на circumvallate; в то время как первые редуцируются в числе, многие, хотя и значительно меньшие, вкусовые почки остаются (Whitehead et al., 1987; Oliver and Whitehead, 1992; Oakley et al., 1993; Sollars et al., 2002; Oakley and Witt, 2004; Guagliardo and Hill, 2007). Напротив, вкусовые почки circumvallate сосочков исчезают в течение первых нескольких недель после денервации (Wong et al., 1994; Seta et al., 1999; Miura et al., 2004), но могут регенерировать после повторного врастания нервов, если нервные оболочки сохранили свою непрерывность (Guth, 1958; Uchida et al., 2003; Miura et al., 2004; Yee et al., 2005). Поскольку дифференцированные вкусовые почки исчезают или уменьшаются в денервированном эпителии, то вполне вероятно, что клетки предшественники также нуждаются в нервных сигналах определенного сорта, чтобы продолжить превращать незрелые клетки во вкусовые почки. Поэтому одной из гипотез является то, что пролиферация базальных кератиноцитов, окружающих вкусовые почки, также должна редуцироваться или останавливаться. Однако, известно лишь единственное исследование по тестированию этой идеи в денервированных грибовидных вкусовых почках, и результаты оказались неубедительны. Наблюдалось лишь незначительное снижение скорости обновления клеток в денервированных вкусовых почках (Oliver and Whitehead, 1992). Но сравнительный анализ пролиферации в более сильно поврежденных circumvallate сосочках мог бы показать, как и когда денервация влияет на предшественники вкусовых клеток. У человека во время хирургических вмешательств в ротовой полости может нечаянно повреждаться как языкоглоточный, так нерв chorda tympani (Gent et al., 2003; Klasser et al., 2008), и chronic otitis media (ушная инфекция) также может приводить к повреждению chorda tympani и к соотв. проблемам восприятия вкуса (Bartoshuk et al., 1996; Gent et al., 2003; Sano et al., 2007; Felix et al., 2009). В каждом таком случае регенерация вкусовых почек из пролиферативного пула скорее всего снижена. Пациенты с раком головы и шеи предоставлют др. указания на природу регенерации вкусовых почек. Эти пациенты получают ежедневные курсы радиотерапии в течение более 6-7 недель, нацеленные на быстро пролиферирующую популяцию опухолевых клеток. Однако при этом постоянно возникает и снижение вкусовой чувствительности у пациентов, что ведет к снижению потребления ghob и потере веса и тем самым к общему снижению качества жизни (Nelson, 1998; Ruo Redda and Allis, 2006; Sandow et al., 2006; Yamashita et al., 2006). При использовании модельных грызунов, облучение также показало потерю вкусовых почек и некоторое нарушение вкусовой чувствительности спустя несколько недель после облучения (Conger and Wells, 1969; Mossman et al., 1979; Nelson, 1998; Yamazaki et al., 2009), то точная клеточно биологическая связь между функцией вкуса и морфологией вкусовых почек и вкусовых нервов не установлена. Пока предложен ряд потенциальных механизмов, наиболее скупой из них, что повторяющаяся (фракционная) радиотерапия вызывает кумулятивное истощение предшественников вкусовых почек, в противоположность прямому повреждению дифференцированных вкусовых почек или афферентных нервных волокон, которые их иннервируют. Эта модель регенерации вкусовых почек согласуется с той, что предложена для облученного поверхностного эпителия. В облученной коже пролиферирующие клетки более чувствительны к вызванным облучением повреждениям ДНК, чем постмитотические клетки и поэтому TA (transit amplifying) пул первоначально повреждается сильнее всего, тогда как значительно меньший комплект слабо делящихся предшественников также уменьшается. Т.о., непосредственно после облучения пролиферация клеток подавляется, поскольку TA и клетки предшественники или пытаются репарировать поврежденную ДНК или, если это безуспешно, подвергаются запрограммированной клеточной гибели. При повторяющихся облучениях увеличивается процент постепенно теряемых TA и клеток предшественников, являющихся их потомками (Dorr et al., 2000; Potten et al., 2002). Применим ли подобный сценарий ко вкусовым почкам предстоит протестировать. Химиотерапия также может вызывать дисфункцию вкусового восприятия у пациентов, получающих лечение при раках, не связанных с головой и шеей (Ovesen et al., 1991; Minakata et al., 2002; Berteretche et al., 2004; Kiewe et al., 2004; Cheng, 2007; Wismer, 2008). И опять быстро делящиеся клетки, подверженные действию лекарств, предположительно предшественники вкусовых почек и TA клетки случайно повреждаются и ведут к истощению клетками вкусовых почек. Наконец, вкусовые ощущения нарушаются при вирусной и бактериальной инфекции (Heald et al., 1998; Bromley, 2000; Pribitkin et al., 2003), возможно из-за воспаления, которое запускается этими патогенами (Wang et al., 2007; Wang et al., 2009; Welge-Luessen, 2009). Фактически, Wang and colleagues (Wang et al., 2007; Wang et al., 2009) показали, что экспериментальная или вирусная инфекция мышей может вызывать специфические interferon реакции во вкусовых почках и в конечном итоге вызывать усиление апоптоза вкусовых клеток. Этот результат указывает на дополнительный уровень регуляции регенерации вкусовых почек, при этом дифференцированные вкусовые клетки являются популяцией мишенью.
Сайт создан в системе
uCoz
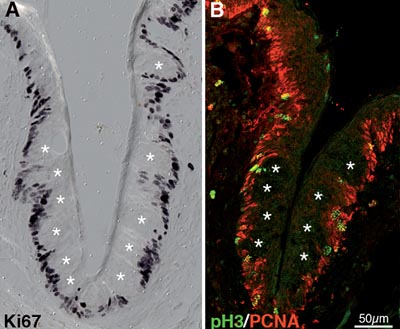 Fig. 1. - Immunostaining with a variety of cell cycle markers reveals that cells within taste buds (white asterisks) of the circumvallate taste papilla are postmitotic. In A and B, only one cleft of the circumvallate papilla is shown in sections taken transverse to the tongue. Taste buds are arrayed in the epithelium lining the cleft, with their apices directed toward the cleft lumen. A. Ki67 marks cells in all phases of the cell cycle, including G1, S, G2 and M, but not resting cells in G0 (Schwarting et al., 1986). Note that basal and perigemmal epithelial cells are immunostained, while intragemmal cells are not. B. PCNA (red) is expressed by cells in all phases of the cell cycle except G1 (Celis et al., 1986), while phosphoHistone3 (pH3; green) is expressed only during M phase (Norbury and Nurse, 1992). Again, immunostained cells are located outside of taste buds (white asterisks), defining the location of the proliferating cells within the circumvallate papilla. The dorsal surface of the tongue is up in both panels. Images courtesy of Ha M. Nguyen.
Fig. 1. - Immunostaining with a variety of cell cycle markers reveals that cells within taste buds (white asterisks) of the circumvallate taste papilla are postmitotic. In A and B, only one cleft of the circumvallate papilla is shown in sections taken transverse to the tongue. Taste buds are arrayed in the epithelium lining the cleft, with their apices directed toward the cleft lumen. A. Ki67 marks cells in all phases of the cell cycle, including G1, S, G2 and M, but not resting cells in G0 (Schwarting et al., 1986). Note that basal and perigemmal epithelial cells are immunostained, while intragemmal cells are not. B. PCNA (red) is expressed by cells in all phases of the cell cycle except G1 (Celis et al., 1986), while phosphoHistone3 (pH3; green) is expressed only during M phase (Norbury and Nurse, 1992). Again, immunostained cells are located outside of taste buds (white asterisks), defining the location of the proliferating cells within the circumvallate papilla. The dorsal surface of the tongue is up in both panels. Images courtesy of Ha M. Nguyen.