Формирование паттерна пальцев обычно интерпретируется в контексте модели градиента морфогена (1, 2). Предполагаемый морфоген Sonic hedgehog (Shh) , исходящий из зоны поляризующей активности (кластера мезодермальных клеток на задней границе зачатка конечности), формирует градиент с максимальными уровнями сзади. Gli3 является основным медиатором передачи сигналов Shh в развитии конечности и полидактилии (2). Поскольку Shh предупреждает преобразование Gli3 в его репрессивную форму (Gli3R), то градиент Shh транслируется в обратный градиент Gli3R (3, 4). Неожиданной находкой стало то, что мышиные Gli3 и Shh;Gli3 нулевые мутанты обнаруживают идентичный фенотип полидактилии конечностей, демонстрируя, что повторяющиеся серии пальцев могут быть сформированы в отсутствие Shh (4, 5). Скорее, чем подтверждение градиентной модели, эти наблюдения согласуются с моделью типа Тьюринга для формирования скрытого рисунка пальцев (6-11) с динамическими взаимодействиями между молекулами активатора и ингибитора, детерминирующими длину волны специфического рисунка, и продуцирующими периодические паттерны пятен или полос. Предполагается, что такой паттерн действует как молекулярный препаттерн хондрогенеза. Согласно одному из предсказаний модели пальцевой промежуток (digit period) или длина волны, предопределяющие объединенную толщину как пальца, так и межпальцевой области, д. быть предметом модуляции при пертурбациях правильных параметров сети генов. Это д. давать аутоподы с пальцами варьирующей толщины и разного количества, которые, кстати, никогда не наблюдались четко.
Хотя основные молекулы механизма самоорганизации остаются неизвестными, потенциальные кандидаты на роль молекулярных модуляторов системы включают Hox гены (10, 12). Дистальные Hoxa и Hoxd гены, как было установлено, влияют на количество пальцев (13), хотя их специфическая роль остается неясной, возможно из-за их разнобойных взаимодействий с путем Shh-Gli3. Эти взаимодействия включают взаимную транскрипционную регуляцию между Hox генами и Shh, а соединение Hoxd12 с Gli3R приводит в результате к блокаде активности репрессора Gli3R (14-16). В целом, эксперименты с избыточностью и потерей функции подтверждают позитивную связь между Hox генами и количеством пальцев (14, 17-22), это также подтверждается с помощью усиления эктопической активности впереди дистальных Hoxd генов при Gli3-зависимой полидактилии (4, 5). Однако мы показали, что комбинированная делеция Hoxd11-13 и Gli3 усиливает Gli3 полидактилию (23), подтверждая вместо этого негативное взаимоотношение между дистальными Hoxd генами и количеством пальцев.
Hoxd11-13;Gli3 мутанты обнаруживают избыток экспрессии Hoxa13, как у Gli3 мутантов (23). Чтобы определить связь между Hoxa13 и количеством пальцев, мы создали Hoxa13;Gli3 мутантов (see supplementary materials and methods). На день эмбриогенеза 12.5 (E12.5), экспрессия Sox9 маркирует пять пальцевых хондрогенных конденсата в контрольных аутоподах и выявляет задержку в дифференцировке в передней мезодерме и полидактилию, типичную для недостатка Gli3 (Fig. 1A) (24). Hoxa13-/-;Gli3XtJ/XtJ обнаруживают от 7 до 8 пальцевых конденсатов в задней мезодерме плюс диффузную экспрессию Sox9 в передней мезодерме, это, скорее всего, соответствует презумптивным пальцевым конденсатам (Fig. 1A). Даже хотя количество пальцев не было в точности установленным, экспрессия Sox9 подтверждает увеличение количества пальцев у Hoxa13-/-;Gli3XtJ/XtJ по сравнению с мутантами Gli3XtJ/XtJ, подтверждая негативный эффект Hoxa13 на количество пальцев, сходное с таковым для Hoxd генов (23).
Hoxa13
-/-;Gli3
XtJ/XtJ конечности также обнаруживают снижение длины волны и бифуркации пальцев (Fig. 1A). Чтобы количественно оценить оба признака, мы анализировали искривление передне-задних (AP) профилей экспрессии
Sox9 на 4-х равноудаленных позициях вдоль проксимо-дистальной (PD) оси в области пальцев
Hoxa13;Gli3
мутантов, показанных на Fig. 1A (Fig. 1B and supplementary materials). Мы определяли средний пальцевой промежуток (digit period) и AP длину каждого профиля (shown in
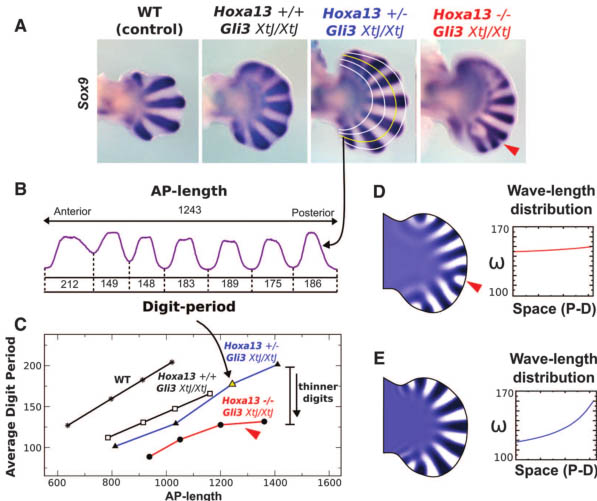 Fig. 1. (A) Expression of Sox9 in E12.5 limbs of the
Hoxa13;Gli3 allelic series. Note the delayed differentiation in the anterior mesoderm in the absence of Gli3.The curved white and yellow lines show the AP profiles used for the analysis of Sox9. The red arrowhead
points to a digit bifurcation. WT, wild type. (B) Sox9
staining intensity along the yellow profile indicated by
the curved arrow. AP length and the period of each
digit (from minimum to minimum) are measured and
shown for Hoxa13+/-;Gli3XtJ/XtJ. (C) Chart showing the
average digit periods versus AP lengths for each
profile and limb. A linear relation is observed in
controls and in the Gli3XtJ/XtJ background for either
the normal or heterozygous dose of Hoxa13,whereas
a flatter relation that correlates with bifurcations (red
arrowhead) is observed in the Hoxa13-/-;Gli3XtJ/XtJ
limbs (red line). The curved arrow marks the yellow
point corresponding to the profile in (B). (D and E)
Two simulations of the reaction-diffusion model
inside an E12.5 Gli3 mutant limb shape. (D) The
activator concentration obtained in the simulation
with a uniform modulation of wavelength w (shown
in the graph) shows digit bifurcation (red arrowhead)
similar to the Hoxa13-/-Gli3XtJ/XtJ mutants. (E) The
simulation result when wavelength is modulated
according to a suitable PD gradient (in this case,a2D
gradient of simulated FGF signaling activity) avoids
bifurcations, because the wavelength increases with
increasing AP length. Limbs shown in all figures are
forelimbs with distal to the right and anterior to the top
Fig. 1. (A) Expression of Sox9 in E12.5 limbs of the
Hoxa13;Gli3 allelic series. Note the delayed differentiation in the anterior mesoderm in the absence of Gli3.The curved white and yellow lines show the AP profiles used for the analysis of Sox9. The red arrowhead
points to a digit bifurcation. WT, wild type. (B) Sox9
staining intensity along the yellow profile indicated by
the curved arrow. AP length and the period of each
digit (from minimum to minimum) are measured and
shown for Hoxa13+/-;Gli3XtJ/XtJ. (C) Chart showing the
average digit periods versus AP lengths for each
profile and limb. A linear relation is observed in
controls and in the Gli3XtJ/XtJ background for either
the normal or heterozygous dose of Hoxa13,whereas
a flatter relation that correlates with bifurcations (red
arrowhead) is observed in the Hoxa13-/-;Gli3XtJ/XtJ
limbs (red line). The curved arrow marks the yellow
point corresponding to the profile in (B). (D and E)
Two simulations of the reaction-diffusion model
inside an E12.5 Gli3 mutant limb shape. (D) The
activator concentration obtained in the simulation
with a uniform modulation of wavelength w (shown
in the graph) shows digit bifurcation (red arrowhead)
similar to the Hoxa13-/-Gli3XtJ/XtJ mutants. (E) The
simulation result when wavelength is modulated
according to a suitable PD gradient (in this case,a2D
gradient of simulated FGF signaling activity) avoids
bifurcations, because the wavelength increases with
increasing AP length. Limbs shown in all figures are
forelimbs with distal to the right and anterior to the top
Fig. 1B for Hoxa13
+/-;Gli3
XtJ/XtJ) и результаты, нанесенные на график (Fig. 1C). В контроле Hoxa13
+/+;Gli3
XtJ/XtJ,
и Hoxa13
+/-;Gli3
XtJ/XtJ мутанты в среднем имеют увеличенный пальцевой промежуток вдоль PD оси, тогда как соотношение между средним пальцевым промежутком (digit period) и AP длиной было постоянным, это указывает, что длина волны в механизме типа Тьюринга масштабирована (scaled) вдоль PD оси, чтобы поддерживать постоянное количество пальцев. Однако это оказывается неверным для
Fig. 2. Representative skeletal phenotypes of new-
borns of the Hoxa13;Hoxd11-13;Gli3 allelic series.
Digit number (indicated for the Gli3XtJ/XtJ condition)
Increases as distal Hox doseis reduced. When only one
Functional copy of Hoxa13 remains (rightcolumn),the
Tip of the digits is connected by acontinuous band of
ossified (red) and cartilaginous (blue) tissue rimming
the distal border of the limb and becoming more
conspicuous as Gli3 copies are removed.
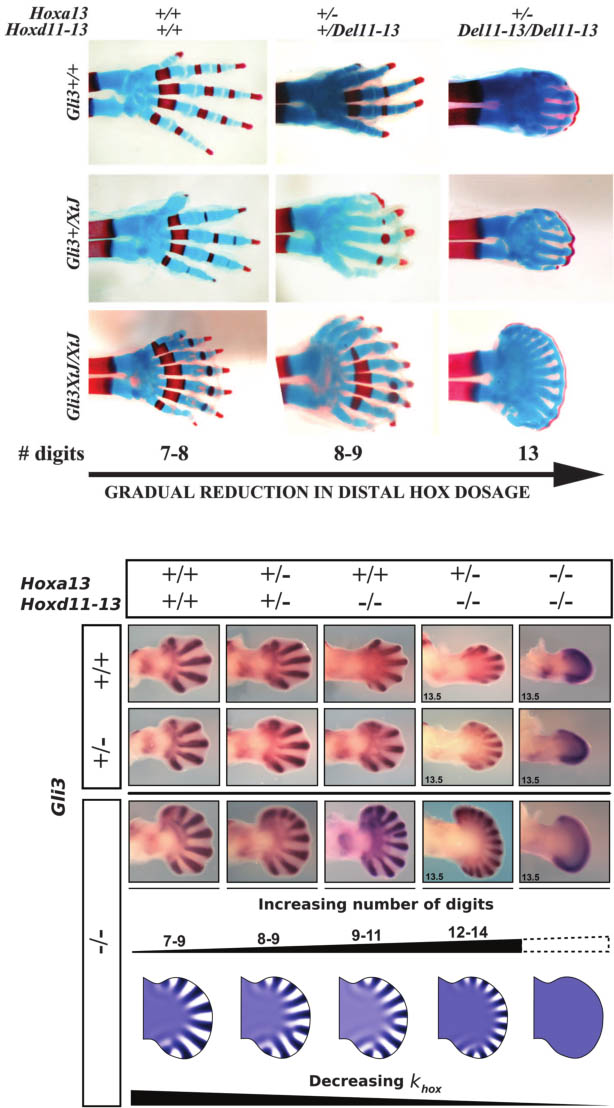
Fig. 3. The phenotypes of triple mutants can be
replicated by the Turing model. (Top) The first three
rows show Sox9 expression at E12.5 and E13.5 for
different combinations of the triple Hoxa13;Hoxd11-
13;Gli3 allelic series. As more Hox are removed, the
general trend shows an increase in digit number and
a decrease in digit thickness. The trend is most
strongly evident in the complete absence of Gli3
(third row). (Bottom) A similar behavior is shown by
the reaction-diffusion simulations, where a decrease
of the PD gradient used to modulate wavelength is
correlated with reduced Hox dose (khox).Additionally,
the model predicts a narrower digital region along
the PD axis, which eventually shrinks to zero, and no
pattern is formed.
Hoxa13
-/-;Gli3
XtJ/XtJ конечностей, где средний пальцевой промежуток сглаживается дистально и в соответствии с Тьюринговым механизмом, появляются бифуркации пальцев, т.к. AP длина увеличивается (Fig. 1A). Наши количественные оценки подтверждают, что формирование нормального паттерна
пальцев использует эффективное PD увеличение пальцевой длины волны, которое, как мы полагаем, должно тщательно контролироваться, чтобы избегать бифуркаций пальцев. Чтобы проверить эту гипотезу мы построили двумерную (2D) модель конечных элементов, базирующуюся
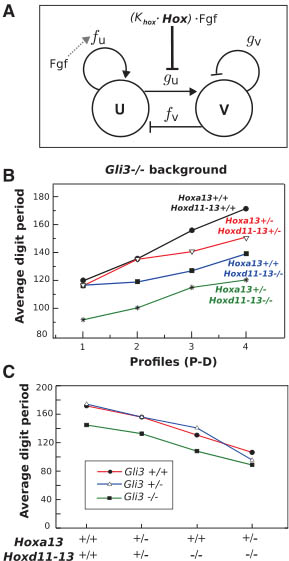 Fig. 4. (A) Schematic representation of the network of a general activator-inhibitor Turing
model. The four reaction kinetic parameters are
shown: fu, fv, gu, and gv. Fgf promotes a PD-graded distribution of the parameter fu to drive stripe orientation (gray dashed arrow). Hox and
Fgf inhibit the parameter gu to increase the wave-
length in a PD-graded manner (bold line). U,
activator; V, inhibitor. (B) Graphs of the average
digit period of the triple mutants with Gli3-/-
background at four equidistant positions along
the PD axis of the digital region. With the exception of Hoxa13+/-;HoxdDel11-13/Del11-13;Gli3XtJ/XtJ,
a clear trend is observed: The PD gradient of wavelength is generally shallower as distal Hox genes are removed. (C) Graphs of averagedigit period (wavelength) versus distal Hox gene dose in the three different Gli3 backgrounds. A smooth positive correlation between Hox gene dose and wavelength is observed in all three cases.
Fig. 4. (A) Schematic representation of the network of a general activator-inhibitor Turing
model. The four reaction kinetic parameters are
shown: fu, fv, gu, and gv. Fgf promotes a PD-graded distribution of the parameter fu to drive stripe orientation (gray dashed arrow). Hox and
Fgf inhibit the parameter gu to increase the wave-
length in a PD-graded manner (bold line). U,
activator; V, inhibitor. (B) Graphs of the average
digit period of the triple mutants with Gli3-/-
background at four equidistant positions along
the PD axis of the digital region. With the exception of Hoxa13+/-;HoxdDel11-13/Del11-13;Gli3XtJ/XtJ,
a clear trend is observed: The PD gradient of wavelength is generally shallower as distal Hox genes are removed. (C) Graphs of averagedigit period (wavelength) versus distal Hox gene dose in the three different Gli3 backgrounds. A smooth positive correlation between Hox gene dose and wavelength is observed in all three cases.
на форме кистевой пластинки Gli3
XtJ/XtJ на ст. E12.5 и смоделировали реакционно-диффузионную систему с помощью масштабирования (scaling) длины волны или униформным (остающимся константным) или градированным способом вдоль PD оси пальцевого региона (Fig. 1, D and E , and supplementary text). Мы использовали характерную для данного класса активатор-ингибитор реакционно-диффузионную систему, употребив минимальные условия, удовлетворяющие нестабильности Тьюринга (see supplementary text) (25). Мы определяли может ли одиночный реакционный параметр модели оказаться эффективной мишенью для модуляции длины волны (figs. S4 and S5) и выбрали эффект активатора на продукцию ингибитора (g
u) как наиболее подходящий (see supplementary text). Наше моделирование показало, что бифуркации пальцев появляются в случае униформности (красная головка стрелки Fig. 1D), тогда как бифуркации не обнаруживаются при использовании масштабирования градированной длины волны (Fig. 1E), воспроизводя мутантные паттерны. Наша модель показывает, что в отсутствие Gli3, генетическое уменьшение Hoxa13 вызывает глобальное уменьшение длины волны (w), тем самым увеличивая количество пальцев, а также вызывая уменьшающийся PD градиент w, это объясняет наблюдаемые бифуркации пальцев. Поскольку отсутствуют доказательства для PD-градированной экспрессии Hox, то простейшей интерпретацией модели является то, что w модулируется как с помощью
Hoxa13 гена (объясняющего глобальное уменьшение длины волны), так и передачи сигналов fibroblast growth factor (FGF) (объясняющей PD градиент), возможно с помощью совместной регуляции одних и тех же генов мишеней (see supplementary text).
Поскольку Hoxa13
-/-;Gli3
XtJ/XtJ двойные мутанты обнаруживают эктопическую переднюю экспрессию
Hoxd12 и Hoxd13, как и
Gli3 мутанты (fig. S1), то оказывается возможным, что увеличение полидактилии обусловлено избытком
Hoxd12 и Hoxd13. Чтобы понять такую функциональную компенсацию и проверить, может ли нарастающая сумма дистальных функциональных Hox генов контролировать количество пальцев, модулированием пальцевого промежутка (digit period), мы получили тройных мутантов, несущих Hoxa13
-, Hoxd
Del11-13 и Gli3
XtJ аллели (Fig. 2). Препараты скелетов новорожденных из аллельной серии тройных мутантов показали варьирующие нарушения формирования паттерна, включая синдактилию (слитые пальцы), брахидактилию
(короткие пальцы), отсутствие суставов, вентральный изгиб пальцев и задержку оссификации. Наиболее выдающимся, однако, была четкая тенденция в направлении увеличения количества пальцев по мере того, как всё больше аллелей дистальных Hox генов удалялось в
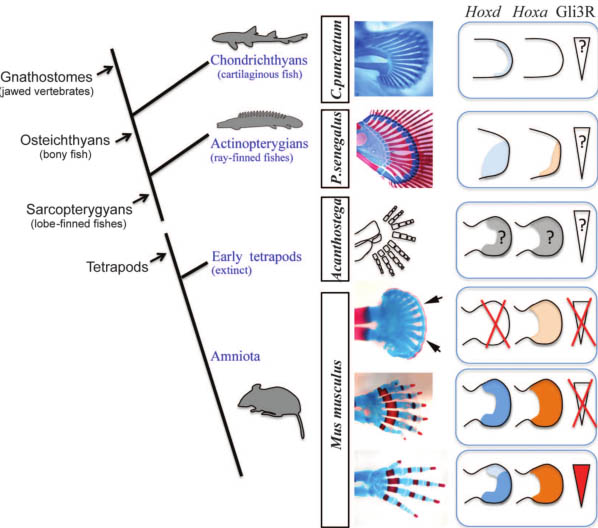
Fig. 5. Vertebrate limb evolution and distal Hox gene function. Aphylogenetic tree
of representative taxa and appendage skeletal patterns is shown, as well as corresponding distal Hox expression [in actinopterygians shown for Polyodon spathula (28)] and Gli3R gradient (when known). Genetically abrogating Shh signaling and reducing distal Hox function in mouse autopods (Hoxa13+/-;HoxdDel11-13/Del11-13;Gli3XtJ/XtJ) reveals ancestral skeletal characteristics shared with the pectoral fins of sharks (Chiloscyllium punctatum) and primitive ray-finned fishes (Polypterus senegalus): numerous, densely packed, and iterative elements, with a distal cartilaginous band corresponding to the distal radials of fish fins (arrows). The periodic pattern of skeletal elements evident in fins and mutant limbs strongly suggests that a self-organizing Turing-type mechanism of chondrogenesis is deeply conserved in vertebrate phylogeny. Our results further indicate that distal Hox gene dose regulates the number and spacing of skeletal elements formed, implicating distal Hox gene regulatory networks as critical drivers of the evolution of the pentadactyl limb.
отсутствие
Gli3 (Fig. 2). Количество пальцев увеличивалось с 7 до 9 (типично для Gli3
XtJ/XtJ) до 8 - 9, когда один функциональный аллель из
Hoxa13 и Hoxd11-13 удалялся; далее до 9 - 11, когда оба Hoxd11-13 аллеля удалялись (23); и, наконец, до 12 - 14 пальцев, когда оставалась только одна копия Hoxa13 (Hoxa13
+/-;Hoxd
Del11-13/Del11-13;Gli3
XtJ/XtJ) (Fig. 2). Гибель в средине беременности
Hoxa13 гомозиготных мутантов пресекает их анализ на этой стадии. Увеличение количества пальцев, когда удаляются дистальные Hox гены, не связано с увеличением AP размера кистевой пластинки (handplate), как это имеет место у одиночных
Gli3 мутантов (26), а скорее связано с уменьшением длины волны, т.к. пальцы были более тонкими и имели узкие промежутки между ними, в то же время оставались регулярно расположенными в пространстве (Figs. 1A and 2). Эти фенотипы не могут быть объяснены с помощью модели позиционной информации, базирующейся на градиенте морфогена.
Чтобы сравнить принципиальные фенотипы серий тройных аллелей с нашей компьютерной моделью, мы анализировали экспрессию Sox9 на ст. E12.5,
когда начинают закладываться хрящевые конденсаты и д. действовать Тьюринговский механизм. Количественная оценка этих мутантов подтвердила, что отсутствует строгая корреляция между прогрессивным увеличением количества пальцев и размером кистевой пластинки, которая вместо этого содержит более тонкие и более тесно упакованные пальцы (Fig. 3). В нашей компьютерной модели как уровни Hox так и передачи сигналов FGF вносят вклад в модуляцию длины волны (Fig. 4A). Т.о., прогрессивное уменьшение глобального вклада Hox генов (khox), приводит в результате к тому, что w градиент становится ниже, но также всё больше уменьшается в глубину (fig. S6), отражая спад градиента длины волны выраженный количественно у мутантов (Fig. 4B). Т.о., модель оказывается способной воспроизводить наблюдаемые усредненные серии Sox9 фенотипов на Gli3XtJ/XtJ фоне-как увеличение количества пальцев, так и более значительную тенденцию к бифуркациям пальцев (Fig. 3 and supplementary text). Хотя наблюдаемое уменьшение длины волны наиболее сильное в отсутствие Gli3, та же самая тенденция четко прослеживается на Gli3+/+ и Gli3+/XtJ фоне (Fig. 4C and supplementary materials). В этих случаях уменьшение длины волны не всегда продуцирует более высокие количества пальцев, поскольку AP ширина также снижается.
Наше исследование подчеркивает онтогенетическую задержку в проявлении Sox9 паттерна (fig. S2) и уменьшение PD ширины региона образования пальцев (fig. S3).Теоретический анализ выявил, что оба признака естественно предсказываются модулированием gu в модели. Задержка формирования паттерна также подкрепляется возможной ролью передачи сигналов FGF в качестве модулятора Turing, т.к. воздействие FGF4 на мезенхиму зачатка конечности, культивируемого в виде микромассы, ускоряло появление паттерна (25). Стимулированный PD градиент передачи сигналов FGF т.о., транслируется в градиент скорости формирования паттерна, а дальнейший теоретический анализ выявил, что это естественно предсказывает прогрессивное PD сужение зоны образования пальцев (fig. S6 and supplementary test). Ниже определенной Hox дозы, пальцевая зона исчезает полностью. В самом деле, у тройных мутантов, не выявляется отдельных пальцевых конденсатов даже на ст. E13.5 (Fig. 3 and fig. S6).
Итак, наша комбинация генетического, количественного анализа и компьютерного моделирования выявляет отсутствующие кусочки доказательства механизма типа Тьюринга в формировании паттерна пальцев. В то время как многочисленные предыдущие мутанты показывали аномальные количества пальцев, отсутствовали доказательства для параметров, которые бы могли гладко настраивать пальцевую длину волны. Наше открытие и анализ усредненной корреляции между количеством дистальных Hox генов и пальцевой длиной волны, предоставили такое доказательство. Кроме того, связь между нарушением регуляции длины волны и появлением бифуркаций пальцев также строго подтверждает роль дистальных Hox генов в качестве модуляторов длины волны прирожденного самоорганизующего типа Тьюринга механизма, ответственного за формирование паттерна пальцев. Наша модель предсказывает, что избыточная экспрессия Hox генов д. увеличивать длину волну пальцев, хотя это нуждается в более тщательном временном исследовании времени оперирования механизма Тьюринга, и может иметь незначительный эффект, если Hox гены нормально экспрессируются на насыщающих уровнях (22). Модель не позволяет сделать предсказания относительно временной последовательности конденсации пальцев вдоль AP оси. Также вероятность бифуркаций может быть снижена у более крайних мутантов (таких как Hoxa13+/-;HoxdDel11-13/Del11-13;Gli3XtJ/XtJ) из-за сужения пальцевого региона. Кроме того, наш анализ подтверждает роль Gli3 в тонкой настройке длины волны, но это наблюдение не столь очевидно, т.к. наблюдается усредненная тенденция в нашей серии дистальных Hox аллелей (Figs. 3 and 4C and supplementary materials).
Наши результаты также позволяют сделать переоценку функции дистальных Hox генов в наиболее важной инновации позвоночных: переход от плавника к конечности. Возникший консенсус указывает, что генетический инструментарий формирования паттерна плавников и конечностей в основном законсервирован и эволюция пальцев управляется за счет накопления регуляторных изменений, контролирующих пространственное и временное развертывание общего инструментария (27-32). Уменьшение количества дистальных Hox генов в отсутствие Gli3, что делает передачу сигналов Shh несоответствующей, в результате происходит потеря пальцев, определяющая характеристики (пятипалость и сегментированную морфологию) и демонстрируются паттерны, напоминающие паттерны эндоскелета у chondrichthyan и базальных actinopterygian плавников (многочисленные, повторяющиеся, плотно упакованные, редко сегментированные элементы) (Fig. 5). Т.о.. наши данные предоставляют доказательства, что родоначальный Тьюринг-подобный механизм формирования паттерна плавников законсервирован у тетрапод и модифицирован за счет осуществления регулярных изменений в эволюции пальцев. В частности, наши данные подтверждают, что равновесие, возникающее в результате перекрестной регуляции между Shh-Gli3 и дистальными Hox генами может приводить к стабилизации состояния пятипалости более чем 360 миллионов лет тому назад.
