Полу-доминантная мутации мышей
Extra-toes (Xt) и
Strong's
Luxoid (Lst), являются мутациями потери функции, нарушающими zincfinger
кодирудщий транскрипционный фактор
Gli3 (Schimmang et al., 1992;
Hui and Joyner, 1993) и aristaless-related гомеобоксный ген
Alx4 (Qu et al., 1998; Takahashi et al., 1998), соотв. В гомозиготном состоянии обе мышиные мутации вызывают плейотропные и летальные врожденные уродства с определенными преаксиальными полидактилиями конечностей. У обоих мутантов возникает передний эктопический сигнальный Sonic Hedgehog (SHH)
центр в дополнение к таковому в задней мезенхиме почки конечности (Masuya et al., 1995; Buscher et al., 1997; Qu et al., 1997). SHH является морфогеном, продуцируемым поляризующей областью (или ZPA) в задней части почки конечности и являющимся важным для контроля формирования переднезаднего паттерна скелета конечности (zeugopod и autopod; rev. Zeller, 2004). Только один передний zeugopodal элемент и палец формируются в конечностях эмбрионов, лишенных функции SHH (Chiang et al 2001; Kraus et al., 2001). Интересно, что полидактиличные фенотипы, вызываемые мутациями
Gli3 и Alx4 отличаются в отношении их зависимости от передачи сигналов SHH. Полидактилия в Gli3 дефицитных конечностях не зависит от SHH, т.к. конечности, лишенные и
Gli3 и Shh имеют полидактилию идентичную таковой у одиночных мутантов
Gli3. Напротив, мутации, вызываемые нарушением
Alx4 зависят от SHH, т.к. только один палец формируется в конечностях, лишенных и
Alx4 и Shh ( te Welscher et al., 2002b). В самом деле, Gli3 действует первым во взаимном антагонистическом взаимодействии с dHand, что, по-видимому, формирует препаттерн мезенхимы почки конечности еще до активации
Shh (te Welscher et al., 2002a). Напротив, Alx4, по-видимому, не функционирует во время этой ранней детерминирующей фазы, стоящей выше формирования паттерна зачатка конечности, обеспечиваемого SHH. Дополнительные существенные различия между двумя мутантами (Qu et al., 1997; Zuniga et al., 1999; te Welscher et al., 2002b) являются, по-видимому, следствием их дифференциальных функций: передняя экспансия обычно ограниченных задними частями доменов экспрессии
Gremlin и 5'Hoxd генов, очевидна с самых ранних стадий зачатка конечности и далее в
Gli3 дефицитных зачатках конечностей. Напротив, небольшие передние эктопические домены экспрессии появляются значительно позднее в
Alx4 дефицитных зачатках конечностей. Более того, Alx4 экспрессия снижается в
Gli3 дефицитных зачатках конечностей, указывая тем самым, что
Gli3 действует генетически выше
Alx4 (te Welscher et al., 2002a). Однако, оставшаяся, более проксимально ограниченная экспрессия
Alx4 в Gli3
-/- мутантных зачатках конечностей, также указывает, что
Alx4 м. функционировать, по крайней мере частично, от
Gli3. Всё это указывает на то, что Gli3, но не
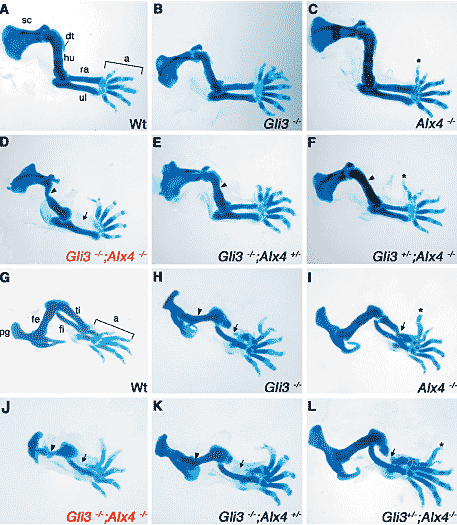 Fig. 1. Skeletal preparations of limbs of single
and compound mutant embryos lacking Gli3
and/or Alx4 at gestational day E14.5. (A-F) forelimbs,
(G-L) hindlimbs, cartilage appears blue as
stained by alcian blue. As ossification is just being
initiated, none or only few ossification centres become
apparent as darker areas (alizarin red). (A)
Wild-type forelimb. (B) Gli3-/- homozygous forelimb.
Note the preaxial polydactyly and loss of digit identity.
(C) Alx4-/- homozygous forelimb. Asterisk indicates
a duplicated preaxial digit 2. (D) Gli3-/-; Alx4-/-
double homozygous forelimb. Arrowhead points to
the malformed humerus. Arrow points to the anterior
zeugopodal region lacking the radius. Note the
duplication of the distal phalange of the most anterior
digit. (E) Gli3-/-; Alx4+/- mutant forelimb, arrowhead
indicates the malformed humerus. The preaxial
polydactyly of the autopod is similar to the one
of a forelimb lacking Gli3 alone. (F) Gli3+/-; Alx4-/-
mutant forelimb. Asterisk indicates the duplicated
preaxial digit 2. Arrowhead points to the humerus
lacking the deltoid tuberosity. Note that the phenotype
of the autopod is identical to the one of a
forelimb lacking Alx4 alone. (G) Wild-type hindlimb.
(H) Gli3-/- homozygous hindlimb. Arrow points to
the rudimentary tibia. (I) Alx4-/- homozygous
hindlimb. Arrow points to the deformed tibia. Asterisk
indicates the duplicated preaxial digit 2. (J)
Gli3-/-; Alx4-/- double homozygous hindlimb. Arrow
points to the region lacking the tibia. Arrowhead
indicates the malformed femur. (K) Gli3-/-; Alx4+/-
mutant hindlimb. The femur (arrowhead) and tibia
(arrow) are affected. (L) Gli3+/-; Alx4-/- mutant
hindlimb. Arrow points to the malformed tibia. Asterisk
points to the duplicated preaxial digit 2. All
panels are oriented with anterior to the top and distal
to the right. a, autopod; dt, deltoid tuberosity; hu,
humerus; ra, radius; sc, scapula; ul, ulna; fe, femur;
fi, fibula; pg, pelvic girdle; ti, tibia.
Alx4
Fig. 1. Skeletal preparations of limbs of single
and compound mutant embryos lacking Gli3
and/or Alx4 at gestational day E14.5. (A-F) forelimbs,
(G-L) hindlimbs, cartilage appears blue as
stained by alcian blue. As ossification is just being
initiated, none or only few ossification centres become
apparent as darker areas (alizarin red). (A)
Wild-type forelimb. (B) Gli3-/- homozygous forelimb.
Note the preaxial polydactyly and loss of digit identity.
(C) Alx4-/- homozygous forelimb. Asterisk indicates
a duplicated preaxial digit 2. (D) Gli3-/-; Alx4-/-
double homozygous forelimb. Arrowhead points to
the malformed humerus. Arrow points to the anterior
zeugopodal region lacking the radius. Note the
duplication of the distal phalange of the most anterior
digit. (E) Gli3-/-; Alx4+/- mutant forelimb, arrowhead
indicates the malformed humerus. The preaxial
polydactyly of the autopod is similar to the one
of a forelimb lacking Gli3 alone. (F) Gli3+/-; Alx4-/-
mutant forelimb. Asterisk indicates the duplicated
preaxial digit 2. Arrowhead points to the humerus
lacking the deltoid tuberosity. Note that the phenotype
of the autopod is identical to the one of a
forelimb lacking Alx4 alone. (G) Wild-type hindlimb.
(H) Gli3-/- homozygous hindlimb. Arrow points to
the rudimentary tibia. (I) Alx4-/- homozygous
hindlimb. Arrow points to the deformed tibia. Asterisk
indicates the duplicated preaxial digit 2. (J)
Gli3-/-; Alx4-/- double homozygous hindlimb. Arrow
points to the region lacking the tibia. Arrowhead
indicates the malformed femur. (K) Gli3-/-; Alx4+/-
mutant hindlimb. The femur (arrowhead) and tibia
(arrow) are affected. (L) Gli3+/-; Alx4-/- mutant
hindlimb. Arrow points to the malformed tibia. Asterisk
points to the duplicated preaxial digit 2. All
panels are oriented with anterior to the top and distal
to the right. a, autopod; dt, deltoid tuberosity; hu,
humerus; ra, radius; sc, scapula; ul, ulna; fe, femur;
fi, fibula; pg, pelvic girdle; ti, tibia.
Alx4, действует первоначально выше SHH, чтобы поляризовать формирующуюся мезенхиму почки конечности (Masuya et al., 1995; Buscher et al., 1997; te Welscher et al., 2002a). Затем SHH обусловленное ингибирование процессинга белка Gli3 продуцирует градиент репрессора Gli3, который как полагают, существенен для формирования передне-заднего паттерна почки конечности (Wang et al., 2000). В ходе дальнейшего развития почки конечности
Alx4 становится критическим для супрессирования передней эктопической экспрессии Shh (Qu et al., 1997). Чтобы выяснить возможные генетические взаимодействия
Gli3 и Alx4, получали
Gli3 и Alx4 компаундных мутантных эмбрионов. После дня беременности 14.5 (E14.5), все генотипы обнаруживались среди выводков в ожидаемой пропорции (Fig. 1 and data not shown), но на E16.5 не обнаруживались двойные гомозиготные эмбрионы
Tabl.1 SKELETAL PHENOTYPES

из-за эмбриональной летальности (data not shown). Скелеты 4-х эмбрионов, лишенные и
Gli3 и Alx4 на ст. E14.5 проанализированы и все конечности двойных гомозигот обладали дефектами скелета, которые отличались от таковых у всех остальных генотипов (Fig. 1, 2 and data not shown; for summary see Table 1). Кроме того, тяжелые черепно-лицевые расщепления в области носа, наблюдались у таких эмбрионов на E14.5 (data not shown).
Skeletal Phenotypes of Forelimbs
Scapula и stylopod в передних конечностях Gli3
-/- ; Alx4
-/- двойных гомозигот уродливы (Fig. 1D; головки стрелок отмечают уродливые humerus), а последний лишен хорошо сформированной deltoid tuberosity. Напротив, humerus у одиночных мутантов
Gli3 (Fig. 1B) и
Alx4 (Fig. 1C) выглядят нормальными, тогда как дополнительная инактивация или
Gli3 или
Alx4 аллеля вызывает легкие уродства humerus (головки стрелок Fig. 1E, F). Эти результаты указывают на дозово-зависимую потребность в этих двух генах во время морфогенеза плечевой кости. В противоположность ко всем остальным генетическим комбинациям (Fig. 1A-C, 1E, F and data not shown), zeugopod передених конечностей Gli3
-/- ; Alx4
-/- двойных гомозигот лишен radius (стрелка, Fig. 1D), тогда как ulna формируется правильно. Наконец, autopod конечностей двойных гомозигот отличен от такового во всех остальных мутантных комбинациях (Fig. 1D and Fig. 2A-F).
Skeletal Phenotypes of Hindlimbs
Как и передние конечности, тазовый пояс и femur задних конечностей двойных Gli3
-/- ; Alx4
-/- гомозигот сильно уродливы (головки стрелок, Fig. 1J). Бедренные кости задних конечностей Gli3
-/- ; Alx4
+/- компаундных мутантов (головки стрелок Fig. 1K) изменены одинаково как и в конечностях Gli3
-/- (головки стрелок Fig. 1H), тогда как они нормальны у эмбрионов, лишенных
Alx4 (Fig. 1I, L). В противоположность передним конечностям tibia отсутствует как в Gli3
-/-; Alx4
-/- так и Gli3
-/-;Alx4
+/- конечностях (стрелки in Figs.
1J, K) и её медиальная часть делетирована и у одиночных Gli3
-/- мутантов(arrow, Fig. 1H) и у Gli3
+/- ; Alx4
-/- компаундных мутантов (arrow, Fig. 1L). Более того, tibia деформирована также у Alx4
-/- одиночных мутантных эмбрионов (Fig. 1I), это указывает на то, что оба гена необходимы и, по-видимому, обладают лишь частично перекрывающимися функциями во время морфогенеза большеберцовой кости. Развитие autopod задних конечностей (Fig. 1G-L) нарушено точно также, как в передних конечностях.
Phenotypes of the Autopod
Дополнительные преаксиальные пальцы формируются в
Alx4-/- конечностях (Fig. 1C, 2C) по сравнению с конечностями дикого типа (Fig. 1A, 2A), независимо от присутствия или обоих (Fig. 1C, 2C; see also Qu et al. 1997) или только одного функционального аллеля
Gli3 (Fig. 1F, 2F). Напротив, autopods
Gli3 дефицитных конечностей обнаруживают тяжелые преаксиальные полидактилии вплоть до 8 пальцев (Fig. 1B, 2B). По сравнению с
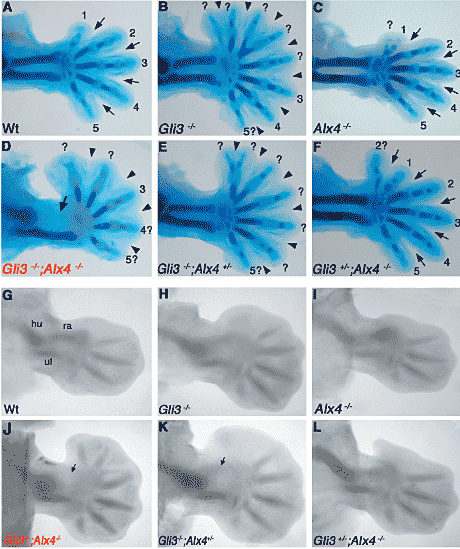 Fig. 2. Skeletal phenotypes forelimbs of different
genotypes at E14.5 and E12.5. (A-F) forelimbs
at gestational day E14.5. (A) Wild-type
control. (B) Gli3 deficient forelimb. Note the preaxial
polydactyly, loss of anterior identities and
webbing. (C) Alx4 deficient forelimb. Note the
additional preaxial digit. (D) Gli3 -/- ; Alx4-/ -
forelimb. Note the lack of the radius (arrow),
duplicated of the distal phalange of the most
anterior digit and general webbing. (E) Gli3-/-;
Alx4+/- forelimb. Note the preaxial polydactyly
and webbing similar to Gli3 -/- forelimbs. (F)
Gli3+/-; Alx4-/- forelimb. Note the additional preaxial digit. Arrows in (A), (C) and (F) indicate the
interdigital areas separating the digits through
apoptosis. Arrowheads in (B), (D) and (E) indicate
webbing in forelimbs of embryos lacking Gli3.
Digits are numbered according to their apparent
identities. Question marks indicate unclear identities.
(G-L) Alcian green staining of the forming
cartilage elements in forelimbs at E12.5. (G-I), (L)
The cartilage primordia of the radius has formed
in (G) wild-type, (H) Gli3-/-, (I) Alx4-/- and (L)
Gli3+/-; Alx4-/- mutant forelimbs. (K) The radius
primordia is only weakly apparent in Gli3-/-;
Alx4+/- mutant forelimbs (arrow). (J) The radius
primordia is lacking in Gli3-/-; Alx4-/- double
mutant forelimbs (arrow). All forelimbs are oriented
with anterior to the top and distal to the
right. hu, humerus; ra, radius; ul, ulna.
Fig. 2. Skeletal phenotypes forelimbs of different
genotypes at E14.5 and E12.5. (A-F) forelimbs
at gestational day E14.5. (A) Wild-type
control. (B) Gli3 deficient forelimb. Note the preaxial
polydactyly, loss of anterior identities and
webbing. (C) Alx4 deficient forelimb. Note the
additional preaxial digit. (D) Gli3 -/- ; Alx4-/ -
forelimb. Note the lack of the radius (arrow),
duplicated of the distal phalange of the most
anterior digit and general webbing. (E) Gli3-/-;
Alx4+/- forelimb. Note the preaxial polydactyly
and webbing similar to Gli3 -/- forelimbs. (F)
Gli3+/-; Alx4-/- forelimb. Note the additional preaxial digit. Arrows in (A), (C) and (F) indicate the
interdigital areas separating the digits through
apoptosis. Arrowheads in (B), (D) and (E) indicate
webbing in forelimbs of embryos lacking Gli3.
Digits are numbered according to their apparent
identities. Question marks indicate unclear identities.
(G-L) Alcian green staining of the forming
cartilage elements in forelimbs at E12.5. (G-I), (L)
The cartilage primordia of the radius has formed
in (G) wild-type, (H) Gli3-/-, (I) Alx4-/- and (L)
Gli3+/-; Alx4-/- mutant forelimbs. (K) The radius
primordia is only weakly apparent in Gli3-/-;
Alx4+/- mutant forelimbs (arrow). (J) The radius
primordia is lacking in Gli3-/-; Alx4-/- double
mutant forelimbs (arrow). All forelimbs are oriented
with anterior to the top and distal to the
right. hu, humerus; ra, radius; ul, ulna.
с заключениями, сделанными в недавних исследованиях (Ahn and Joyner, 2004; Harfe et al., 2004), задние три пальца сохраняют свои качественные особенности, тогда как 5 наиболее пальцев лишены характерных особенностей (знаки вопроса Fig. 2B). Довольно неожиданно полная инактивация
Alx4 и Gli3 (Fig. 1D and Fig. 2D) уменьшает количество пальцев с 8 (Fig. 2B, E) до 5 или 6 (Fig. 1D, 2D, 2J). Задние три пальца сохраняют, по крайней мере, частично задние характеристики (как и у Gli3
-/- ;
compare Fig. 2D to 2B), тогда как передние два пальца не обнаруживают определенных характеристик, чтобы их идентифицировать. Эти результаты показывают, что тяжелые фенотипы преаксиальных полидактилий у
Gli3 дефицитных эмбрионов (Fig. 1B, 2B) зависят от функции Alx4. Т.к. паттерн пальцев со 2 по 5 формируется под влиянием SHH-обусловленного градированного ингибирования образования репрессора Gli3, то самый передний палец 1 и передний zeugopod (radius и tibia) специфицируются с помощью др. ещё неизвестного механизма (rev. Zeller, 2004). Результаты нашего исследования показывают, что взаимодействие транскрипционного фактора Alx4 с Gli3 (repressor) белком может участвовать в спецификации наиболее передних скелетных элементов конечности (digit 1, radius и
tibia). Подтверждает это предположение характерное удвоение преаксиального пальца, наблюдаемое в передених конечностях, лишенных
Alx4, но не
Gli3 (Fig.
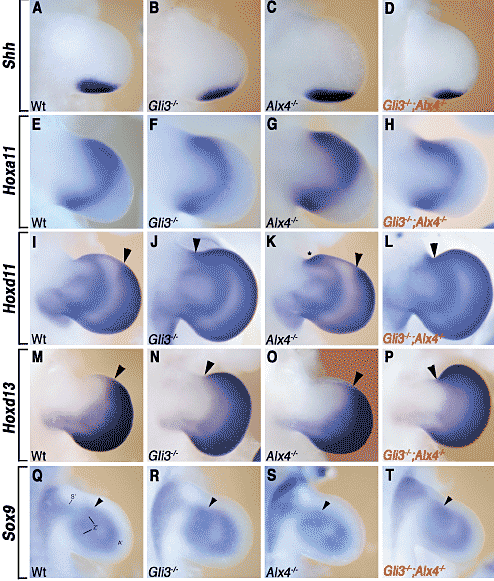 Fig. 3. Molecular analysis of key regulators
of limb bud patterning. (A-D) Shh
expression in wild-type and mutant forelimb
buds (E10.75). At this stage, there are
no significant differences in Shh expression
between wild-type (A), single (B,C)
and double homozygous mutant limb buds
(D). Anterior ectopic Shh expression is
detected much later in mutant limb buds
(data not shown; Buscher et al., 1997;
Masuya et al., 1995; Qu et al., 1997). (E-H)
Expression of Hoxa11 in wild-type and
mutant forelimb buds (E11.5). Note that
the Hoxa11 expression domain in Gli3-/-
; Alx4-/- limb buds (H) is identical to the
one in Gli3-/- limb buds (F). (I-L) Hoxd11
expression in wild-type and mutant forelimb
buds (E11.75). Again the Hoxd11
expression domain in Gli3-/-; Alx4-/- (L)
and Gli3-/- limb buds (J) are similar. (M-P)
Hoxd13 expression in the distal limb mesenchyme
of wild-type and mutant limb
buds (E11.75). The Hoxd13 expression in
Gli3-/-; Alx4-/- limb buds (P) again resembles
the one of Gli3-/- limb buds (N).
Arrowheads in (I-P) indicate the anterior
boundaries of the distal (autopod) expression
domains. Asterisk in (K) indicates the
ectopic anterior domain of Hoxd11 expression
(Qu et al., 1997). (Q-T) Sox9
expression in wt and mutant limb buds
(E11). Sox9 marks pre-cartilaginous condensations
of mesenchymal cells. No significant
differences are apparent. Arrowheads
point to the approximate position of
the condensations giving rise to the primordia
of the radius. S': prospective
stylopod; Z': prospective zeugopod; A':
prospective autopod. All limb buds are
oriented with anterior to the top and distal
to the right
Fig. 3. Molecular analysis of key regulators
of limb bud patterning. (A-D) Shh
expression in wild-type and mutant forelimb
buds (E10.75). At this stage, there are
no significant differences in Shh expression
between wild-type (A), single (B,C)
and double homozygous mutant limb buds
(D). Anterior ectopic Shh expression is
detected much later in mutant limb buds
(data not shown; Buscher et al., 1997;
Masuya et al., 1995; Qu et al., 1997). (E-H)
Expression of Hoxa11 in wild-type and
mutant forelimb buds (E11.5). Note that
the Hoxa11 expression domain in Gli3-/-
; Alx4-/- limb buds (H) is identical to the
one in Gli3-/- limb buds (F). (I-L) Hoxd11
expression in wild-type and mutant forelimb
buds (E11.75). Again the Hoxd11
expression domain in Gli3-/-; Alx4-/- (L)
and Gli3-/- limb buds (J) are similar. (M-P)
Hoxd13 expression in the distal limb mesenchyme
of wild-type and mutant limb
buds (E11.75). The Hoxd13 expression in
Gli3-/-; Alx4-/- limb buds (P) again resembles
the one of Gli3-/- limb buds (N).
Arrowheads in (I-P) indicate the anterior
boundaries of the distal (autopod) expression
domains. Asterisk in (K) indicates the
ectopic anterior domain of Hoxd11 expression
(Qu et al., 1997). (Q-T) Sox9
expression in wt and mutant limb buds
(E11). Sox9 marks pre-cartilaginous condensations
of mesenchymal cells. No significant
differences are apparent. Arrowheads
point to the approximate position of
the condensations giving rise to the primordia
of the radius. S': prospective
stylopod; Z': prospective zeugopod; A':
prospective autopod. All limb buds are
oriented with anterior to the top and distal
to the right
1C, F and Fig. 2C, F) отсутствующее также в передних конечностях у большинства двойных мутантов (Fig. 1D, 2D). Однако, наиболее дистальная фаланга наиболее переднего пальца часто удвоена (Fig. 1D, 2D). В противоположность инактивации обоих
Alx4 аллелей (стрелки, Fig. 2C, F), потеря функции Gli3 вызывает образование межпальцевой кожной складки (головки стрелок, Fig. 2B, E) в результате блокирования межпальцевого апоптоза (Macias et al., 1999). Эта межпальцевая кожная складка не меняется в результате конкурентного удаления
Alx4 в конечностях двойных гомозигот (Fig. 2D), это подчеркивает неперекрывающуюся и важную функцию Gli3 в межпальцевом апоптозе.
Phenotypes of the Zeugopod
Наиболее тяжелыми дополнительными дефектами скелета конечностей у эмбрионов, лишенных и
Gli3 и Alx4 по сравнению с одиночными гомозиготными мутантами является потеря переднего zeugopodal
элемента (radius и tibia, Fig. 1). Анализ формирования паттерна зачатков конечностей на стадии (E12.5, Fig. 2G-L) показал, что хрящевая модель, дающая radius не образуется у двойных гомозигот (стрелка, Fig. 2J). Неожиданно при анализе более поздних стадий (Fig. 1E, 2E), зачаток radius также менее очевиден у Gli3
-/- ; Alx4
+/- компаундных мутантов на этой стадии (стрелка, Fig. 2K), чем в конечностях дикого типа (Fig. 2G) и у др. генотипов (Fig. 2H, I, L and data not shown). Эти результаты указывают на то, что образование radius задерживается в конечностях, лишенных
Gli3 и одной копии
Alx4 (Fig. 2K), но оно нагоняется во время последующего развития (Fig. 1E, 2E) в противоположность аплазии в передних конечностей двойных мутантов (Fig. 1D, 2D, 2J).
Normal Expression of Molecular Markers of Limb Bud Patterning
Т.к. в особенности формирование передних скелетных элементов конечностей нарушено у двойных гомозиготных мутантов, то анализировалась экспрессия соотв. ключевых регуляторов. Shh продолжает экспрессироваться нормально в задней мезенхиме одиночных и двойных гомозиготных зачатках конечностей (Fig. 3A-D). Не наблюдается преждевременной передней эктопической экспрессии Shh в зачатках конечностей двойных гомозигот (Fig. 3D) как это имеет место в зачатках конечностей одиночных мутантов (Masuya et al., 1995; Buscher et al., 1997; Qu et al., 1997). Генетический анализ установил, что паралоги гены Hoxa11 и Hoxd11 взаимодействуют, чтобы специфицировать ulna и radius (Davis et al., 1995), а проксимальная неправильная экспрессия Hoxd13 ингибирует морфогенез zeugopod (Goff and Tabin, 1997). Следовательно, эти три Hox гена являются хорошими кандидатами для детекции альтераций, затрагивающих формирование паттерна zeugopod. Однако, экспрессия транскриптов ни Hoxa11, ни Hoxd11 (Fig. 3H, L) достоверно не меняется в зачатках передних конечностей Gli3-/- ; Alx4-/- двойных гомозигот по сравнению с Gli3 дефицитными конечностями
(Fig. 3F, 3J). Более того, не выявлено достоверных изменений в экспрессии Hoxd13 в зачатках конечностей Gli3-/-; Alx4-/- двойных гомозигот (Fig. 3P) по сравнению с одиночными мутантными и зачатками конечностей дикого типа (Fig. 3M-O). Следовательно, отсутствие radius не обнаруживает параллелизма с соотв. изменениями в паттернах экспрессии этих трех Нох генов во время стадии формирования паттерна зачатков конечностей. В частности, передние границы доменов экспрессии Hoxd11 и Hoxd13 в дистальной мезенхиме зачатков конечностей у Gli3-/- ; Alx4-/- эмбрионов (головки стрелок, Fig. 3L, P) идентичны с таковыми в зачатках конечностей Gli3-/- (головки стрелок, Fig. 3J, N) и отличаются существенно от таковых в зачатках конечностей Alx4-/- и дикого типа (головки стрелок, Fig. 3I, K, M, O). Эти результаты показывают, что Gli3, но не Alx4, необходим, чтобы регулировать передние границы доменов экспрессии Hoxd11 и Hoxd13. Установление этих доменов экспрессии регулируется с помощью Gli3 уже во время начала развития зачатков конечностей (Zuniga and Zeller, 1999). Наконец, анализ транскрипционного фактора Sox9, экспрессируемый сконденсированными мезенхимными клетками, которая предетерминирует форму элементов хряща конечностей (Wright et al., 1995), показал, что мезенхимные конденсаты индуцируются. по-видимому, нормально в зачатках конечностей Gli3-/- ; Alx4-/- двойных гомозигот (Fig. 3T; compare to Fig. 3Q-S).
Показанные на Figure 3 результаты говорят о том, что события формирования паттерна, управляющие инициацией мезенхимных конденсатов и хрящевых моделей, в частности зачат ков передних конечностей, происходят в той же самой степени в зачатках конечностей Gli3-/- ; Alx4-/- и Gli3 -/-. Данное исследование выявляет перекрывающиеся функции Gli3 и Alx4 в формировании radius, тогда как укорочение (но не полная потеря) tibia было описано ранее в конечностях Alx4-/- и Gli3-/- одиночных мутантов (Johnson, 1967; Qu et al., 1998). В зачатках передних конечностей, лишенных Fgf4 и Fgf8, скелетные предшественники, формирующие zeugopodal конденсаты, редуцированы, что наиболее вероятно обусловливает непосредственно потерю radius (Sun et al., 2002). Напротив, кажущееся нормальным распределение Sox9 в зачатках конечностей Gli3-/- ; Alx4-/- двойных мутантов указывает на то, что популяция скелетных предшественников не редуцирована и индукция мезенхимных конденсатов происходит нормально. Дефекты, затрагивающие zeugopod также обнаруживаются и у разнообразных др. мутаций, в частности у тех, которые затранивют 5'Hoxd и 5'Hoxa гены (Zakany and Duboule,
1999). Напр., мышиная мутация Ulnaless (Ul ) затрагивает цис-регуляцию 5'Hoxd генов (Spitz et al., 2003). Эктопическая проксимальная экспрессия Hoxd13 вместе с редуцированной экспрессией Hoxd11 вызывает тяжелую редукцию обоиз zeugopod скелетных элементов в конечностях мутантных Ul эмбрионов (Herault et al., 1997; Peichel et al.,
1997). Более того, Chen et al. (2004) установили, что репрессор Gli3 взаимодействует непосредственно с Hoxd белками, чтобы обеспечивать спецификацию пальцев. Следовательно, можно предположить, что специфические генетические взаимодействия Gli3 с Alx4 участвуют в морфогенезе зачатков конечностей на уровне регуляции и функции Hox белков скорее, чем их экспрессии.
Сайт создан в системе
uCoz THIJS DRENTH, PASCAL TEWELSCHER, AIMEE ZUNIGA and ROLF ZELLER
THIJS DRENTH, PASCAL TEWELSCHER, AIMEE ZUNIGA and ROLF ZELLER