Некоторые гены участвуют в сборке или функционировании ресничек и могут обладать дополнительными функциями, не связанными с ресничками, так что будучи мутантными они вызывают комбинацию связанных с ресничками и не связанных с ресничками симптомов. Напр., huntingtin-interacting protein Hippi/ IFT57, который, по-видимому, играет роль как в цилиогенезе, так и апоптозе (Houde et al.. 2006). Болезни или мутации,изменяющие функцию Hippi, могут т.о. влиять как на симметрию тела (связанную с ресничками) , так и на апоптическую стрессовую реакцию в головном мозге (преимущественно не связано с ресничками). Сходным образом многие белки клеточной полярности необходимы для цилиогенеза (Schermer et al.. 2006: Fan et al.. 2007), так что мутации таких генов могут вызывать дефекты ресничек в комбинации с др. связанными с полярносттью фенотипами. Даже белки, играющие очень специфические роли в функции ресничек, могут обладать дополнительными функциями. Напр., intraflagellar transport (IFT) мотор KIF3 участвует также в аксональном транспорте (Kondo et al.. 1994). Сходным образом GAS11 является компонентом dynein регуляторного комплекса, белкового комплекса, присоединенного к наружным дублетам, который регулирует активность dynein плеч, но он также локализуется в аппарате Гольджи, где он может обеспечивать взаимодействия между Гольджи и микротрубочками (Colantonio et al.. 2006).
Исследования на модельных организмах, таких как
C. elegans показали, что не все реснички являются одинаковыми в отношении аппарата, необходимого для их строительства и поддержки. Известно, что в некоторых типах клеток
 Figure 1. The anatomy of the cilium. Cilia are arrays of nine microtubule doublets that extend from the basal body (a modified cenfriole) and push out an extension of the plasma membrane (called the ciliary membrane). Some cilia also contain a pair of singlet microtubules in the middle called the central pair that is involved in regulating motility. Motile power is provided by two sets of dynein arms: the inner and outer arms. Motility is regulated by radial spokes that interact with the central pair. Defects in any of these structures can lead to ciliary diseases, although the precise symptoms that arise appear to depend on which structure is altered.
Figure 1. The anatomy of the cilium. Cilia are arrays of nine microtubule doublets that extend from the basal body (a modified cenfriole) and push out an extension of the plasma membrane (called the ciliary membrane). Some cilia also contain a pair of singlet microtubules in the middle called the central pair that is involved in regulating motility. Motile power is provided by two sets of dynein arms: the inner and outer arms. Motility is regulated by radial spokes that interact with the central pair. Defects in any of these structures can lead to ciliary diseases, although the precise symptoms that arise appear to depend on which structure is altered.
микротрубочки ресничек подвергаются постоянному обмену (Stephens. 1997), тогда как в др. аксонемные микротрубочки, по-видимому, значительно менее динамичны. Следовательно, степень, с которой аппарат цилиогенеза, включая IFT (-see Seholey on p. 23 of this issue), необходим в этих разных типах ресничек, д. быть безусловно отличной, более динамичные реснички нуждаются в более высокой эффективности постоянной сборки. Такие реснички д. первыми подвергаться болезненным мутациям, которые частично редуцируют IFT. Следовательно, гипоморфные аллели генов цилиогенеза могут вызывать дефекты только в субнаборе ресничек, давая фенотип, который отличен от такового у нулевых мутантов.
Существуют также четко отличающиеся потребности для сборки ресничек в разных типах клеток в одном и том же организме. У C. elegans разные сенсорные реснички имеют драматически отличающуюся морфологию, при этом некоторые имеют сложную разветвленную или веерообразную структуру, а др. имеют более каноническую форму цилиндрической реснички. Помимо этих структурных различий разные типы ресничек отличаются также по их потребности в разных частях аппарата IFT. Два разных кинезина, kinesin-II и OSM-3.. обычно кооперируют, чтобы построить сенсорную ресничку, при этом OSM-3 специфически необходим для построения дистальной половины реснички, которая содержит только singlet микротрубочки и преимущественно специализируется для выполнения сенсорной функции (Ou et al.. 2005). Интересно, что в ресничках AWB нейронов два моторных белка более не связаны в отличие от др. типов проанализированных ресничек. OSM-3 более не нужен, чтобы строить дистальный сегмент (Mukhopadhyay et al.. 2007). Даже в более экстремальном случае некоторые структуры ресничек, такие как жгутики спермиев Drosophila melanogaster не нуждаются в IFT для всей их сборки (Han et al.. 2003). У диатомовых отсутствие ретроградного IFT мотора, вместе с IFT complex A и Bardet-Biedl syndrome proteins (Seholey. 200S) в геноме указывает на то, что эти части IFT систем являются необязательными в некоторых случаях (Merchant et al., 2007). Если сходная изменчивость потребности в IFT обнаруживается и между разными клетками и типами тканей у людей, то можно представить, что генетические дефекты в разных компонентах аппарата IFT могут иметь более тяжелые дефекты ресничек в некоторых типах клеток, чем в др.
Драматические различия в механизме цилиогенеза наблюдаются в мультиреснитчатом эпителии. В противоположность первичной ресничке в большинстве клеток, реснички в клетках воздушных путей и эпендимуса закреплены с помощью базальных телец, которые формируются de novo в крупных сферических образованиях, наз. deuterosomes (Dirksen. 1991). Дефекты генов, специфичных для deuterosome, могут приводить к дефектам ресничек только специфического мультиреснитчатого эпителия, не оказывая какого-либо эффекта на сенсорные первичные реснички.
Др. важным соображением является относительное время потери гена в разных тканях. Цилиопатии с началом заболевания у взрослых возникают в результате спонтанной потери гетерозиготности в цилиарном гене у пациентов, несущих мутантным один из аллелей. Этот второй удар д. происходить спустя длительное время после эмбриогенеза: т.о. дефекты развития, такие как полидактилия не д. встречаться. Время начала цилиопатии может быть изучено, используя индуцибельные Cre-опосредованные нокаутные аллели (Garcia-Gonzalez et al.. 2007).
Defects affecting different ultrastructurai modules of cilia
Реснички имеют модулярную организацию на ультраструктурном уровне (Fig. 1), и индивидуальные структурные модули (напр., центральная пара, динеиновые плечи и радиальные спицы) выполняют разные функции. Напр., dynein рукава или радиальные спицы необходимы для подвижных, но не для сенсорных ресничек, тогда как каналы мембраны ресничек могут быть необходимы для восприятия, но не для движения
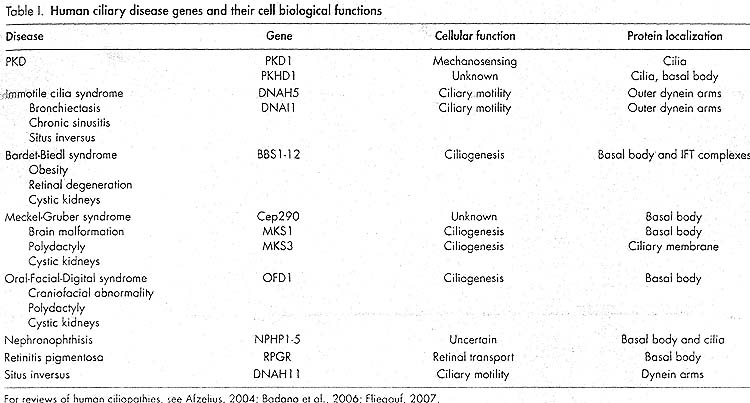
Т.о., воспроизведение определенной болезни может быть связано с нарушениями некоторых специфических субнаборов структурных и функциональных свойств ресничек, оставляя др. интактными.
Этот эффект четко наблюдается при primary ciliary dyskinesia (PCD), которая также известна как синдром неподвижных ресничек. PCD обычно связана с дефектами dynein плеч, радиальных спиц или центральной пары (которые являются компонентами аппарата движения). Дефекты в таких структурах, как ожидается, не д. затрагивать передачи сигналов: напр., таких какие обычно наблюдаются при полидактилии или др. дефектах передачи сигналов hedgehog во время развития, у пациентов с PCD. Сходным образом пациенты с PCD не д. страдать кистозом почек, ожирением или дегенерацией сетчатки, т.к. эти симптомы возникают благодаря дефектам в структуре ресничек, которые не связаны с подвижностью.
Можно ещё дальше подразделять случаи PCD на те, что затрагивают различные структуры подвижности. Основные симптомы PCD, а именно дефекты в очищении слизистой, обнаруживаются при каждом дефекте структур подвижности. Др. симптом наиболее часто ассоциированный с неподвижными ресничками это гидроцефалия, отражающая роль ресничек эпендимных клеток в движении спинномозговой жидкости. Гидроцефалия, по-видимому, специфически ассоциирует с дефектами центральной пары микротрубочек, т.к. это было продемонстрировано на hydin мутантных мышах, которые характеризуются гидроцефалией и отсутствием центральной пары микротрубочек (Davy and Robinson, 2003: Lechtreck and Witman. 2007). Третий симптом неподвижных ресничек это situs inversus, инверсия обычно лево-правосторонней асимметрии внутренних органов. Situs inversus появляется из-за подвижности первичных ресничек в узелке млекопитающих, генерирующей левонаправленный ток жидкости, который нарушает лево-правостороннюю симметрию во время развития. Однако реснички узелка, по-видимому, лишены центральной пары микротрубочек, по крайней мере, в раннем развитии и в соответствии с этим отсутствием центральной пары на этой стадии, мутации, которые ведут к потере центральной пары микротрубочек (напр., hydin мутанты), обычно не вызывают situs inversus.
Определенные субструктуры ресничек могут также играть определенную и отдельную роль в передаче сигналов. обеспечиваемой ресничками. Мутация hennin мышей вызывает изменение пространственной распространенности передачи сигналов hedgehog во время развития (Caspary et al., 2007). Интересно, что эффект на hedgehog может осуществляться посредством альтераций структуры самих ресничек (Caspary et al.. 2007), делая B трубочку наружных дублетов дефектной с разрывом в точке, в которой она встречается с A трубочкой, обращенной внутрь реснички. Протофиламента B трубочки, которая взаимодействует с а трубочкой в этом регионе имеет необычный вид на электронных микрофотографиях и может даже не состоять из tubulin (for review see Linck and Stephens. 2007), поэтому было бы интересно выяснить функциональные последствия потери этого очень необычного структурного элемента. Мутанты fleer рыбок данио также имеют дефект в соединении В трубочек с А трубочками, очевидно в результате потери fleer-специфической polyglutamylation тубулина. Однако в отличие от фенотипа hennin у мутантов fleer разрыв между А и В трубочками находится на др. A-B соединении, обращенном к мембране реснички (Pathak et al., 2007). Эти мутанты обнаруживают дефекты в подвижности ресничек, но не имеют драматического hedgehog фенотипа. Трудно сравнивать мутантов мышей и рыбок данио непосредственно из-за крупных материнских накоплений у эмбрионов рыбок, которые могут замаскировать ранние онтогенетические фенотипы. Поэтому эти результаты предполагают возможность, что специфические протофиламенты В трубочек могут играть специфические роли в передаче сигналов.
Если структуры, участвующие в подвижности также участвуют и в передаче сигналов во время развития. то следует ожидать, что пациенты с дефектами этих структур скорее всего не будут доживать до рождения. Этим можно объяснить, почему мутации наружного плеча dynein также часто выступают в качестве причины PCD у пациентов (Hornet et al.. 2006) ? т.к. мутации наружных дублетов или центральной пары могут в конце концов приводить к более тяжелым симптомам.
Мембрана ресничек является др. компонентом, в котором располагаются функциональные молекулы. Широкий круг рецепторов и каналов локализован в этой специализированной мембране, которая является критической для функции реснички. Напр., временные receptor potential каналы активно перемещаются вдоль мембраны ресничек (Qin et al.. 2005), хотя значение этого перемещения для передачи сигналов остается неизвестным. Одним из наиболее охарактеризованных мембранных каналов ресничек является сенсор механических стрессов, состоящий из polycystins, мутации которого вызывают polycystic kidney disease 1 (PKD1) и PKD2, аутосомно доминантные PKD. Эти белки локализуются в ресничке (Barr et al.. 2001: Yoder et al., 2002) и, как полагают, наделяют реснички способностью ощущать ток жидкости в почках. PKD пациенты с дефектами этих белков не обнаруживают др. дефектов ресничек, указывая тем самым, что PKD белки не обязательны для цилиогенеза или др. функций ресничек. Передача сигналов ресничками в конечном итоге передает информацию в цитоплазму и ядро. Дефекты в определенных нижестоящих сигнальных путях могут воспроизводить эффект потери ресничек в тех типах клеток, которые имеют отношение к этим путям, тогда как потери этой функции ресничек в др. типах клеток не наблюдается. в
Дефекты в трафике и биогенеза мембран ресничек также могут приводить к болезням. Bardet-Biedl syndrome белковый комплекс взаимодействует с guanine nucleotide exchange фактором для Rab5, который сам по себе, по-видимому, регулирует рекрутирование мембраны в ресничку (Nachury et al., 2007). Дефекты доставки компонентов мембраны реснички могут давать легкие изменения функции ресничек.
Наконец. можно представить, что произойдет, когда ресничка полностью нормальна в терминах структуры и состава, но просто находится в неправильном месте и в несоответствующее время. Подвижные реснички д. быть соотв. ориентированы, чтобы направлять ток в правильном направлении. Реснички также д. быть на соответ. поверхности клетки (напр., обращены в просвет протока), чтобы правильно воспринимать окружение. Одним из наиболее драматических типов неправильно локализации ресничек является описанный у пациентов является направление ресничек во внутриклеточные вакуоли скорее, чем на клеточную поверхность (Hagiwara et al.. 2000). В этих случаях можно увидеть, что сенсорные реснички продолжают функционировать, но вместо восприятия окружения, для которого клетка предназначена (напр.,просвета протока), они ощущают срееду внутри вакуоли. Она д. отличаться совершенно от нормальной внеклеточной среды и может несоответствующим образом активировать сигнальные пути, базирующиеся на ресничках, с серьезными негативными последствиями для функции клеток.
Future directions
The bottom line is that cilia are complex, and details of the complexity vary from one cell type to another. To understand the full spectrum of ciliary diseases, therefore, we must learn much more about cell type-specific differences in cilia. A key first step would be to use proteomics to identify differences in the composition of different types of cilia. Another important step would be the development of simple assays for ciliary function and structure that can be applied in a range of tissues, both in animal models and in samples from human patients, to understand how different mutations affect different types of cilia.
Сайт создан в системе
uCoz