Геном человека содержит миллионы энхансеров7. Эти участки ДНК действуют как переключатели, регулирующие, где и когда экспрессируются все гены, кодирующие белки в нашем геноме. Таким образом, энхансеры кодируют инструкции для тканеспецифичной экспрессии генов и, следовательно, успешного развития, гомеостаза взрослого организма и целостности клеток8. Большинство SNV, связанных с фенотипическими изменениями и заболеваниями, расположены в энхансерах1-3. Определение того, какие SNVs в энхансере способствуют развитию заболевания, представляет собой огромную проблему, поскольку эти причинные варианты часто находятся в море инертных вариантов1-3,9. Наша неспособность точно определить причинные варианты энхансеров является узким местом в установлении связи между генотипом и фенотипом. Здесь мы используем механистические и обобщающие принципы, управляющие энхансерами, для предсказания причинных вариантов энхансеров. Такой подход может обеспечить систематические и масштабируемые методы, которые используют весь потенциал геномных данных для улучшения здоровья человека.
Для изучения взаимосвязи между последовательностью энхансеров и фенотипами мы сосредоточились на энхансере ZRS4. Этот энхансер регулирует экспрессию Shh (SHH у человека) в задней части развивающихся зачатков конечностей в области, известной как зона поляризующей активности (ZPA), и имеет решающее значение для развития конечностей и пальцев у позвоночных животных, таких как цыплята, мыши и человек10-12. Этот энхансер длиной около 800 п.н. высоко консервативен по последовательности между мышами и людьми, и у обоих видов он расположен на расстоянии почти 1 Мб от промотора Shh4. Хотя ZRS является одним из наиболее хорошо изученных энхансеров, остается неясным, как он кодирует экспрессию генов и как SNV вызывают фенотипические отклонения. Более 30 SNV в ZRS, обнаруженных у позвоночных, связаны с полидактилией и другими дефектами конечностей, такими как тибиальная гемимелия (укорочение голени) (дополнительная таблица 1). Несколько человеческих семей и мышей имеют одинаковые SNV в ZRS и демонстрируют схожие фенотипы, что свидетельствует об устойчивости фенотипов полидактилии в разных видах и на генетических фонах (Дополнительная таблица 1). Высокая степень сохранения формирования паттерна пальцев и последовательности ZRS у мышей и людей делает мышь отличной системой для изучения генетических основ полидактилии. Исследования, анализирующие влияние SNVs на энхансерную активность ZRS у мышей, позволяют предположить, что полидактилия связана с эктопической энхансерной активностью в зачатке передней конечности, обусловленной усилением функции (GOF). Восемь человеческих SNV, ассоциированных с полидактилией, были протестированы в эндогенном локусе мыши, и исследования показали, что четыре из них вызывают GOF экспрессии Shh в зачатке передней конечности и дополнительные пальцы5,13 (дополнительная таблица 2). Однако механизмы, с помощью которых эти SNV изменяют функцию энхансера, плохо изучены5,13,14.
Redundant low-affinity ETS sites regulate the ZRS
ZRS регулируется комбинацией транскрипционных факторов, включая HAND2, HOX, ETV4, ETV5, ETS-1 и GABPa13,15-18. Пять аннотированных сайтов, известных как ETS1-ETS5, участвуют в транскрипционной активации Shh из ZRS и связываются с транскрипционными факторами ETS-1 и GABPa15. Как ETS-1, так и GAPBa активируются ниже по течению от сигнализации фактора роста фибробластов (FGF) из апикального эктодермального гребня19. Удаление всех пяти сайтов ETS приводит к полной потере энхансерной активности в ZPA при тестировании репортерных последовательностей у мышей15. Удаление отдельных сайтов не влияет на экспрессию, однако удаление комбинаций этих сайтов приводит к значительному снижению экспрессии в ZPA15. Эти результаты показывают, что все пять сайтов ETS (ETS1-ETS5) необходимы для активации экспрессии Shh в ZPA. Новый принцип регуляции энхансеров, в том числе регулируемых ETS, заключается в использовании субоптимально-аффинных сайтов связывания (также известных как низкоаффинные или субмаксимальные сайты связывания) для кодирования тканевой специфичности энхансера20-22. Этот принцип изучался в основном у беспозвоночных20-23.
Чтобы выяснить, придерживается ли ZRS позвоночных также этого принципа регуляции, мы измерили относительную аффинность (сродство) пяти сайтов ETS (ETS1-ETS5), используя данные микрочипов связывания белков (PBM) для мышиного транскрипционного фактора ETS-1 (ссылки 24,25). PBM измеряет сродство связывания всех возможных 8-мерных последовательностей для интересующего транскрипционного фактора, чтобы обеспечить прямое измерение связывания25. Относительная аффинность рассчитывается путем сравнения сигнала всех 8-меров с сигналом самого высоко-аффинного сайта, который имеет оценку 1.00 или 100%. Специфичность связывания ДНК ETS-1 и других факторов транскрипции ETS класса I консервативна у мышей и людей (расширенные данные рис. 1). Поэтому сродство связывания, измеренное для ETS-1, передает сродство связывания других факторов транскрипции I класса ETS, которые экспрессируются в почке конечности и которые также могут связываться с этим локусом, например GABPa15,17,24. Хотя PBM измеряется in vitro, относительная аффинность, определенная с помощью PBM, показывает сильную корреляцию с интенсивностью пиков при иммунопреципитации хроматина (ChIP) in vivo в нескольких наборах данных (расширенные данные, рис. 2).
Пять ранее аннотированных и функционально подтвержденных сайтов ETS (ETS1-ETS5)
15 в ZRS имеют неоптимальное сродство связывания, варьирующее от 0,26 до 0,39 относительно консенсуса (рис. 1а). Мы идентифицировали в общей сложности 19 предполагаемых ETS-сайтов в ZRS человека и 18 в ZRS мыши, все из которых имеют субоптимальное сродство. Пятнадцать из этих сайтов являются консервативными по расположению и сродству между человеком и мышью (расширенные данные рис. 3а). Один из этих сохраненных сайтов - недавно идентифицированный сайт ETS-A, который имеет чрезвычайно низкое сродство 0,15. Мы подтвердили, что эта последовательность ETS-A связывает транскрипционный фактор ETS-1 с помощью анализа сдвига электрофоретической подвижности (EMSA) (рис. 1а и расширенные данные рис. 4).
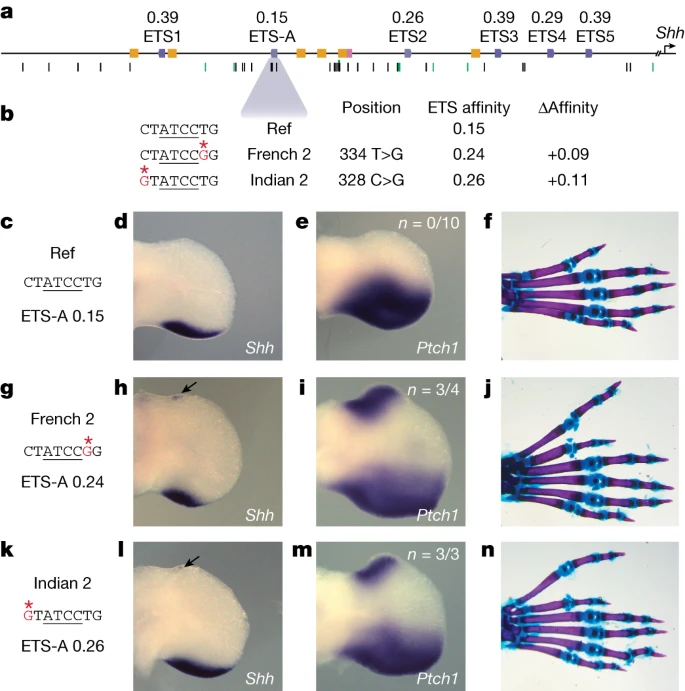
Fig. 1:
An ETS-A site in the ZRS enhancer contains two human variants that are associated with polydactyly, both of which subtly increase ETS binding affinity.
a, The human ZRS contains five known and functionally validated ETS sites, ETS1–ETS5, all of which have suboptimal affinity. We identify a new site, ETS-A, which has a relative affinity of 0.15. Six HOX sites (yellow) and one HAND2 site (pink) have also been previously identified. Thirty-one SNVs associated with polydactyly are found in humans (black bars), and SNVs are also found in other species such as cats, mice and chicks (green bars). b, Two human SNVs associated with polydactyly, denoted French 2 and Indian 2, occur within the ETS-A site. Both SNVs lead to a subtle increase in relative affinity of ETS-A to 0.24 and 0.26 respectively. c–f, The ETS-A sequence in a reference (Ref) mouse (c) drives the expression of Shh (d) and Ptch1 (e) restricted to the posterior domain of the developing limb bud in E11.75 and E12.0 embryos, respectively, as shown by in situ hybridization. f. Skeletal staining shows a WT mouse hindlimb with normal digit morphology. g–j, The French 2 SNV (g) drives the ectopic expression of Shh (h) and Ptch1 (i) in the anterior limb bud of homozygous embryos (arrow) in addition to the normal domain of posterior expression. j, A mouse hindlimb homozygous for French 2 has an extra triphalangeal thumb. k–n, The Indian 2 SNV (k) drives the ectopic expression of Shh (l) and Ptch1 (m) in homozygous embryos (arrow). n, A hindlimb from an Indian 2 homozygous mouse has an extra triphalangeal thumb. We did not calculate n for Shh because the expression is highly dynamic and thus hard to accurately capture; instead, we calculate the n of Ptch1 as a readout of Shh. Human polydactyly SNVs subtly increase affinity
Сайт ETS-A находится в области ZRS, которая полностью сохраняется между мышами и людьми (расширенные данные, рис. 3б). Два варианта человека, ассоциированные с полидактилией, - French 2 (334 T>G) и Indian 2 (328 C>G) - расположены в сайте ETS-A (рис. 1b). Вариант French 2 встречается в семье с преаксиальной полидактилией; он является неполно пенетрантным, поскольку три из четырех членов семьи с этим вариантом имеют лишний большой палец6. Идентифицирован только один человек с индийским вариантом 2, у которого наблюдается преаксиальная полидактилия5. Примечательно, что оба человеческих варианта вызывают сходное незначительное увеличение относительного сродства к сайту ETS-A с 0,15 у референса до 0,24 у French 2 и 0,26 у Indian 2. EMSA подтверждает, что оба варианта связываются с ETS-1 сильнее, чем ETS-A дикого типа (WT) (расширенные данные рис. 4). Мы предположили, что это небольшое 1,6-кратное увеличение относительного сродства к сайту ETS-A может быть причиной полидактилии. Хотя оба варианта были изучены с помощью репортерных анализов LacZ у мышей, эти анализы показывают, что только вариант French 2 изменяет активность энхансера5. Ни один из этих человеческих вариантов не был изучен в эндогенном локусе ZRS, и механизм, с помощью которого они изменяют экспрессию генов, неизвестен. Поэтому сначала мы попытались определить, проявляется ли у мышей с вариантами French 2 и Indian 2 эктопическая экспрессия Shh и преаксиальная полидактилия.
У гомозиготных мышей French 2 и Indian 2 наблюдается эктопическая экспрессия Shh в передней части задней конечности на эмбриональный день (E)11.75 (рис. 1h,l). Область эктопической экспрессии Shh крошечная и очень динамичная. Поэтому мы также рассмотрели Ptch1, прямую мишень Shh, которая обычно используется в качестве индикатора передачи сигналов Shh13,26. Ptch1 эктопически экспрессируется у гомозигот French 2 и Indian 2 E12.0 (рис. 1i,m). Мы не наблюдали эктопической экспрессии Shh или Ptch1 в передней конечности на ст. E11.75 или E12.0, что, вероятно, объясняется различиями в регуляции экспрессии Shh в передней и задней конечности27. У гетерозиготных и гомозиготных мышей French 2 и Indian 2 наблюдается преаксиальная полидактилия задних конечностей, что свидетельствует о том, что очень небольшая и преходящая эктопическая экспрессия Shh может оказывать сильное влияние на количество пальцев (рис. 1j,n). У человека полидактилия чаще всего возникает на передних конечностях, а у мышей - на задних5,6,13 (Дополнительная таблица 1). Вероятно, это связано с различиями в спецификации передних и задних конечностей у этих двух видов28,29. Дополнительный передний палец в задних конечностях мыши напоминает дополнительный трехфаланговый палец, наблюдаемый при ортологичных врожденных пороках развития у человека, и мы называем его трехфаланговым пальцем ноги. Таким образом, оба варианта являются причинно-следственными для полидактилии и фенокопируют наблюдаемый фенотип человека.
Affinity-optimizing SNVs cause polydactyly
Варианты French 2 и Indian 2 имеют одинаковый фенотип, что позволяет предположить, что механизмом, обуславливающим полидактилию, может быть одно и то же тонкое увеличение сродства сайта ETS-A. Чтобы проверить это предположение, мы создали еще две линии мышей путем манипуляций внутри сайта ETS-A. Первая линия мышей, Syn 0.25, содержит синтетически созданный сайт ETS-A с аффинностью 0.25 - такой же, как у вариантов French 2 и Indian 2, но с другим изменением последовательности (рис. 2а). Далее мы проверили связывание ETS-1 с каждой из этих последовательностей ETS-A с аффинностью примерно 0,25 с помощью EMSA и не обнаружили существенной разницы в связывании ETS-1 с последовательностями French 2, Indian 2 или Syn 0,25 (расширенные данные, рис. 4). Возможно, что любое нарушение последовательности ETS-A может привести к фенотипическому отклонению. Чтобы показать, что это не так, мы также создали линию мышей, которая, как мы предсказывали, не будет иметь никакого влияния на фенотип. Мы создали мышь с потерей функции (LOF), у которой сайт связывания ETS-A был уничтожен путем удаления важнейшего нуклеотида, необходимого для связывания (рис. 2e и расширенные данные рис. 4). Из-за избыточности сайтов ETS в ZRS
15 мы предсказали, что потеря сайта ETS-A не повлияет на экспрессию
Shh или развитие конечностей.

Fig. 2:
Synthetic changes to the ETS-A site that create a 0.25-affinity site and a LOF site cause predicted phenotypes.
a–d, The Syn 0.25 ETS-A site (a), which has an affinity of 0.25, drives the ectopic expression of Shh (b) and Ptch1 (c) in the anterior limb bud, in addition to the normal domain of expression in the posterior limb bud. d, Skeletal staining of a homozygous mouse hindlimb shows an extra triphalangeal thumb. e–h, The LOF ETS-A site (e) drives the normal expression of Shh (f) and Ptch1 (g) in the posterior limb bud. h, Homozygous mice have normal digit morphology. We did not calculate n for Shh because the expression is highly dynamic and thus hard to accurately capture; instead, we calculate the n of Ptch1 as a readout of Shh.
У мышей, содержащих сайт Syn 0.25 ETS-A, наблюдается эктопическая экспрессия Shh и Ptch1 в зачатке передней конечности в возрасте E11.75 и E12.0, соответственно, что отражает картину экспрессии, наблюдаемую у мышей French 2 и Indian 2 (рис. 2b,c). У мышей Syn 0.25 также наблюдается преаксиальная полидактилия задних конечностей (рис. 2d), что позволяет предположить, что фенотип обусловлен изменением аффинности, а не изменением последовательности. Как и предполагалось, мыши с мутацией LOF не демонстрируют эктопической экспрессии Shh или Ptch1 (рис. 2f,g) и имеют нормальную морфологию конечностей (рис. 2h). В совокупности эти исследования показывают, что GOF-увеличение аффинности ETS-A в энхансере ZRS является патогенным, в то время как LOF-вариант непатогенен. Эта работа демонстрирует нашу способность успешно предсказывать связь между генотипом и фенотипом для вариантов последовательности в сайте ETS-A.
Мы предположили, что если все три варианта, оптимизирующие сродство к ETS-A, имеют одинаковый механизм действия, то пенетрантность, латеральность и тяжесть полидактилии должны быть сопоставимы между тремя линиями. Фенотипирование мышей проводилось вслепую по отношению к генотипу. Мыши French 2, Indian 2 и Syn 0.25 с одинаковым сродством к сайту ETS-A имеют сходную пенетрантность и латеральность у гетерозигот, при этом полидактилия чаще всего возникает на правой задней конечности (рис. 3а). У гомозигот фенотип проявляется двусторонне и пенетрантность полидактилии выше, чем у гетерозигот. Хотя все мыши в этом исследовании были выведены в идентичных условиях и имеют одинаковый генетический фон, в каждой линии наблюдается разброс фенотипов (тяжести) пальцев (рис. 3б). При этом распределение фенотипов пальцев по линиям идентично. У гетерозигот наиболее часто встречается двухфаланговый или трехфаланговый дополнительный палец, в то время как у гомозигот наиболее часто встречается трехфаланговая дополнительный палец. Таким образом, эти три линии мышей с одинаковым увеличением сродства демонстрируют одинаковую пенетрантность, латеральность и тяжесть заболевания без статистических различий.
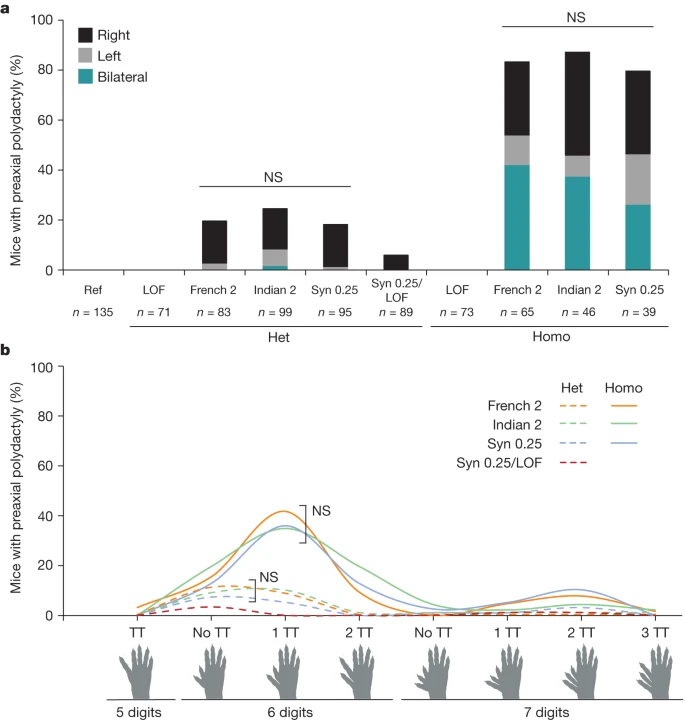
Fig. 3:
All mice with the approximately 0.25-affinity ETS-A are indistinguishable in terms of the penetrance, laterality and severity of polydactyly.
a, Penetrance and laterality of phenotypes seen in French 2, Indian 2 and Syn 0.25 heterozygous (Het) and homozygous (Homo) mice. There is no significant difference in penetrance and laterality between heterozygotes and between homozygotes of French 2, Indian 2 and Syn 0.25 mice (Fisher’s exact test). Polydactyly occurs more frequently on the right hindlimb in both heterozygotes and homozygotes. In Syn 0.25/LOF mice, polydactyly is less penetrant than in heterozygous Syn 0.25 mice (Syn 0.25/WT). NS, not significant. b, Polydactyly phenotypes seen in French 2, Indian 2 and Syn 0.25 heterozygotes (dashed lines) and homozygotes (solid lines), as well as in Syn 0.25/LOF mice. There is no significant difference in the phenotypes seen between the French 2, Indian 2 and Syn 0.25 lines (Fisher’s exact test). TT, triphalangeal toe. Biorender.com was used to generate images in this figure.
Далее мы хотели выяснить, действительно ли сайт ETS-A является функциональным, что довольно сложно из-за избыточности сайтов в энхансере
15. Если сайт WT ETS-A вносит свой вклад в активность энхансера, то пенетрантность полидактилии у мышей, содержащих одну копию аллеля ETS-A Syn 0.25 и одну копию сайта WT ETS-A, должна быть выше, чем у мышей с одним аллелем ETS-A Syn 0.25 и одним аллелем LOF ETS-A. Возникновение и выраженность фенотипов у мышей ETS-A Syn 0.25/LOF ETS-A значительно снижены по сравнению с мышами ETS-A Syn 0.25/WT ETS-A (рис. 3), что свидетельствует о том, что 0.15-аффинный сайт WT ETS-A вносит свой вклад в энхансерную активность и фенотипы.

Fig. 4:
A greater increase in affinity at the ETS-A site causes more severe and penetrant polydactyly and long-bone defects.
a, The ETS-A reference (Ref) sequence. b, In situ hybridization of Shh in WT hindlimb at E11.75 shows a domain of expression in the posterior of the limb bud. c, Skeletal staining of a WT mouse hindlimb showing five digits. d, WT forelimb showing five digits e, WT hindlimb focused on tibia and fibula. f, The Syn 0.52 ETS-A sequence. g, In situ hybridization of Shh in a Syn 0.52 homozygous mouse hindlimb bud at E11.75. Ectopic expression in the anterior of the limb bud is indicated by an arrow. h, Skeletal staining of a homozygous Syn 0.52 ETS-A mouse hindlimb showing six digits. The toe and the extra digit are both triphalangeal. i, Homozygous Syn 0.52 mouse forelimb showing six digits. Both the thumb and the extra digit are triphalangeal. j, Hindlimb focused on the tibia and fibula in a Syn 0.52 ETS-A homozygous mouse. The tibia is severely shortened. The hindlimbs in e,j were imaged with the same magnification. See Extended Data Fig. 6 for more details.
Наш комплексный анализ фенотипов 795 трансгенных мышей показал, что все три варианта, увеличивающие сродство к ETS-A примерно до 0,25, имеют неразличимые фенотипы у гетерозигот и гомозигот. Это убедительно свидетельствует о том, что тонкая оптимизация сродства этого сайта ETS-A является механизмом, обуславливающим полидактилию. Несмотря на растущее признание роли низкоаффинных сайтов в энхансерах, сайты с аффинностью 0,15 и даже 0,25 обычно игнорируются5. Однако в данном случае мы видим, что сайт с аффинностью 0,15 является функциональным, а сайт с аффинностью 0,25 не только функционален, но и достаточен для нарушения нормального развития конечностей, это указывает на то, что незначительное увеличение количества сайтов с низкой аффинностью может быть патогенным.
Predicting penetrance and severity
Убедившись, что небольшое увеличение аффинности может вызывать дефекты развития, мы задались вопросом, может ли степень изменения аффинности предсказать пенетрантность и тяжесть фенотипов. Такое механистическое понимание может оказаться ценным для диагностики и лечения. Чтобы проверить эту гипотезу, мы создали линию мыши с 0,52-аффинным сайтом ETS-A (рис. 4f). EMSA подтверждает, что эта последовательность связывает ETS-1 сильнее, чем WT ETS-A или 0,25 ETS-A сайты (расширенные данные рис. 4).
Гомозиготы French 2, Indian 2 и Syn 0.25 имеют небольшую величину эктопической экспрессии Shh в задней конечности, а сайт Syn 0.52, как и предсказывалось, вызывает формирование большого домена экспрессии Shh и Ptch1 в передней части задней конечности, а также эктопическую экспрессию в передней конечности (рис. 4g и расширенные данные рис. 5g,i). В соответствии с этими паттернами экспрессии у мышей Syn 0.52 наблюдается полидактилия как передних, так и задних конечностей, в то время как у мышей с аффинностью 0.25 полидактилия наблюдается только на задних конечностях (рис. 4h,i). Более того, у мышей Syn 0.52 полидактилия полностью проявляется в задних конечности как у гетерозигот, так и у гомозигот, и почти полностью проявляется в передних конечностях (расширенные данные рис. 6а). Полидактилия у мышей Syn 0.52 чаще всего двусторонняя как у гетерозигот, так и у гомозигот, тогда как у 0.25 ETS-A аффинных мышей преобладает односторонний фенотип. Большинство мышей Syn 0.52 имеют шесть трехфаланговых пальцев, но некоторые из них имеют семь или даже восемь пальцев; это более серьезное явление, чем полидактилия, наблюдаемая у мышей, родственных 0.25 ETS-A (расширенные данные рис. 6c). Синдактилия также встречается чаще у мышей Syn 0.52, чем у мышей French 2, Indian 2 и Syn 0.25. Кроме того, у мышей Syn 0.52 наблюдаются дефекты длинных костей. Укорочение большеберцовой кости или гемимелия большеберцовой кости - состояние, наблюдаемое у людей13 , - встречается у 95 % гомозигот Syn 0.52 (рис. 4j и расширенные данные рис. 6b). Таким образом, как и предсказывалось, большее увеличение сродства приводит к более глубокому нарушению и тяжелым фенотипам. Это дает основания полагать, что увеличение сродства может быть использовано для прогнозирования тяжести и пенетрантности фенотипов.
Affinity-optimizing SNVs prevalent across the ZRS
Далее мы хотели проверить, может ли наша способность предсказывать причинные варианты энхансеров распространяться на другие сайты ETS в пределах ZRS. Семнадцать SNVs вызывают увеличение сродства к ETS по меньшей мере в 1,6 раза; это изменение в 2 раза происходит во French 2, Indian 2 и Syn 0,25 (рис. 5a). Чтобы выяснить, вызывают ли эти варианты усиление функции, мы проанализировали данные опубликованных репортерных анализов ZRS. В одном из этих анализов, в котором проверяется влияние мутагенеза 2% последовательности энхансера ZRS человека на экспрессию в зачатках конечностей мыши5 , мы обнаружили, что варианты энхансера, содержащие оптимизирующие сродство SNV ETS, значительно обогащены при экспрессии GOF. Однако внутри каждого энхансера происходит около 16 п.н. изменений, поэтому мы не можем однозначно приписать экспрессию GOF отдельному SNV (рис. 7 расширенных данных).
Fig. 5: Affinity-optimizing SNVs drive GOF expression in the ZRS and IFN? enhanceosome...
Наш анализ данных этих двух исследований мутагенеза ZRS5,30 и углубленное изучение вариантов в сайте ETS-A показывают, что оптимизирующие аффинность SNV в четырех различных сайтах ETS в ZRS могут вызывать экспрессию гена GOF.
Чтобы проверить, распространяются ли наши выводы за пределы ETS на другие сайты связывания транскрипционных факторов (TFBS) в ZRS, мы провели поиск HOX-оптимизирующих SNV, связанных с полидактилией. HOXA13 и HOXD13 экспрессируются в дистальной части зачатка конечности, и изменения в их экспрессии могут влиять на развитие пальцев и конечностей31. SNV Dutch 2 (165 A>G32) увеличивает сродство к HOXA13 и HOXD13. EMSA подтверждает, что SNV Dutch 2 связывает эти HOX-белки сильнее, чем последовательность WT, что позволяет предположить, что увеличение связывания HOX лежит в основе фенотипа полидактилии Dutch 2 (расширенные данные рис. 8a,b и дополнительная таблица 3).
The enhanceosome contains affinity-optimizing SNVs
Enhanceosome интерферона-и (IFNβ) - это хорошо охарактеризованный энхансер, который включает экспрессию гена IFNβ в качестве иммунного ответа на вирусную инфекцию33,34. Для активности энхансера необходимы IRF-связывающие сайты35. Мы проанализировали результаты MPRA-анализов, в которых мутировала каждая пара оснований в этом энхансере36. SNVs, которые увеличивают аффинность сайтов связывания IRF с изменением в 1,5 раза, значительно обогащают экспрессию GOF по сравнению со всеми другими SNV в MPRA, в то время как нет значительного обогащения экспрессии GOF в SNV внутри сайтов IRF, которые не изменяют аффинность связывания IRF (рис. 5c). Один из этих SNV, оптимизирующих сродство к IRF, был независимо протестирован в другом анализе и также показал экспрессию GOF35. Эти результаты служат дополнительным доказательством общей роли аффинно-оптимизирующих SNV в экспрессии GOF в другом энхансере, регулируемом разными транскрипционными факторами и активном в разных клетках. В отличие от энхансера ZRS, enhanceosome является избыточным энхансером37,38. Таким образом, в контексте репортерных анализов принцип оптимизации сродства применим к двум классическим энхансерам, примерам избыточного и нередуцированного энхансера.
Other transcription factors and disease enhancers
Чтобы изучить роль вариантов, оптимизирующих аффинность, в других энхансерах, мы рассмотрели MPRA-анализ 11 энхансеров, связанных с различными заболеваниями30 (рис. 9 расширенных данных). Для оценки влияния каждого основания на активность энхансера использовался насыщающий мутагенез. Каждый MPRA проводился в клеточной линии, соответствующей каждому конкретному энхансеру30. Поскольку FGF-сигнализация важна для многих типов клеток, а нарушения FGF-сигнализации связаны с различными заболеваниями, мы не отфильтровывали энхансеры, реагирующие на ETS, а просто просмотрели все 11 энхансеров. Медиана сродства к ETS в этих энхансерах составляет 0,12. Энхансеры с SNV, оптимизирующими аффинность ETS (~1,6 раза), значительно обогащены для экспрессии GOF, в то время как SNV, которые встречаются в сайтах ETS, но не изменяют их аффинность, не значительно обогащены в GOF энхансерной активности (рис. 6a). Существует множество SNV, которые незначительно увеличивают аффинность связывания и связаны с активностью энхансера GOF, это указывает на то, что небольшое увеличение аффинности связывания может способствовать экспрессии гена GOF в широком диапазоне энхансеров.
Fig. 6: Affinity-optimizing SNVs drive GOF expression in a wide variety of disease-associated enhancers. ..
Мы также рассмотрели AP-1, транскрипционный фактор, который участвует во многих клеточных процессах, включая дифференциацию и пролиферацию клеток39. Поскольку AP-1 является широко распространенным транскрипционным фактором, мы снова не стали отфильтровывать энхансеры, которые, как известно, регулируют AP-1, а просто просмотрели все 11 энхансеров. Аналогичным образом мы обнаружили, что SNV в MPRA-анализе, которые увеличивают сродство к AP-1 (~1,5 раза), приводят к экспрессии генов GOF, в то время как SNV в сайтах AP-1, которые не изменяют сродство, не значительно обогащают экспрессию GOF (рис. 6b).
Чтобы проверить, насколько наши выводы применимы к другим наборам данных, мы проанализировали MPRA-скрин, в котором проверялись регуляторные элементы лимфобластоидов и варианты внутри этих элементов, выявленные в исследовании экспрессии локусов количественных признаков (eQTL)40. В этом исследовании были изучены геномы и транскриптомы лимфобластоидов 446 индивидов йоруба и европейского происхождения, а также индивидов из проекта "1000 геномов "41. Геномы и мРНК из этих лимфобластоидных клеток были проанализированы, чтобы соотнести геномные вариации с изменениями в экспрессии генов. Для тестирования в MPRA были выбраны наиболее ассоциированные геномные варианты или варианты, находящиеся в неравновесии по сцеплению с этими вариантами eQTL. В общей сложности с помощью MPRA было измерено влияние более 3 000 SNV в той же клеточной линии, что и при исследовании eQTL. Как и предсказывалось, мы наблюдаем значительное обогащение энхансерами GOF активности в пределах SNV, оптимизирующих аффинность как для ETS, так и для AP-1. SNV, встречающиеся в ETS или AP-1, которые не изменяют аффинность энхансеров, не обогащены активностью GOF (рис. 6c,d). Вместе взятые, эти анализы двух ортогональных наборов данных MPRA показывают, что для двух разных транскрипционных факторов, в огромном разнообразии контекстов, сайты SNVs, оптимизирующие аффинность, являются общим механизмом, приводящим к экспрессии генов GOF в репортерных анализах.
MPRA позволяют изучать влияние вариантов на экспрессию в контексте репортерного анализа, в то время как eQTL определяет влияние вариантов на экспрессию целевого гена в контексте генома. Из семи SNV, оптимизирующих сродство к ETS GOF, выявленных в MPRA лимфобластоидов, пять являются значимыми eQTL. Все пять eQTL ассоциированы с увеличением экспрессии генов-мишеней (расширенные данные рис. 10a). Таким образом, увеличение экспрессии репортеров, наблюдаемое при MPRA, коррелирует с экспрессией целевого гена в эндогенном локусе. Действительно, проанализировав сигнал eQTL всех 2 663 вариантов eQTL в исследовании40 , мы обнаружили, что SNVs, оптимизирующие сродство к ETS, значительно обогащают GOF экспрессии целевых генов (расширенные данные, рис. 10b). Напротив, варианты eQTL в сайтах ETS, которые не изменяют сродство, не вызывают значительного обогащения в экспрессии GOF для генов-мишеней.
В масштабах всего генома оптимизирующие сродство ETS SNV в eQTL демонстрируют значительное обогащение экспрессии целевых генов (положительные значения бета), тогда как SNV, не изменяющие сродство сайтов ETS в энхансерах, не показывают обогащения экспрессии целевых генов41 (Extended Data Fig. 10c). Более того, с помощью набора данных eQTL мы обнаружили, что более высокие изменения сложения ETS показывают более значительное обогащение экспрессии генов-мишеней GOF (расширенные данные, рис. 10d). Мы также видим такое обогащение для AP-1 (Extended Data Fig. 10e). Хотя не все изменения в экспрессии, обнаруженные при анализе eQTL, являются прямыми, наблюдаемое нами обогащение свидетельствует о том, что оптимизирующие сродства SNV управляют GOF-экспрессией генов-мишеней в эндогенном локусе и что эта GOF-активность не буферизируется в эндогенном контексте.
Regulatory principles predict causal SNVs
Энхансеры изобилуют вариациями, поэтому предсказание того, какие SNV являются каузальными, представляет собой серьезную проблему для связи генотипа с фенотипом. В наборе данных ZRS MPRA 14,5% всех SNV приводят к экспрессии GOF, в то время как 36% оптимизирующих сродство SNV приводят к экспрессии гена GOF. Поиск сродство-оптимизирующих SNV ETS в ZRS увеличивает нашу способность находить причинные варианты усилителей GOF в 2,5 раза, если рассматривать только данные MPRA (рис. 6e). В MPRA пропущены два варианта GOF, которые мы выявили в этом исследовании (French 2 и Indian 2). Таким образом, при учете всех известных аффинных SNV в ZRS, 6/11 или 55% аффинных SNV в ETS приводят к экспрессии GOF, что увеличивает нашу способность находить случайные варианты в наборе данных в 3,8 раза (рис. 6e). Действительно, поиск SNV, повышающих аффинность ETS, AP-1 и IRF, улучшает нашу способность находить причинные варианты GOF от двух раз до семи раз (рис. 6e). Поиск SNV, оптимизирующих сродство, является эффективным и простым методом выявления предполагаемых каузальных вариантов энхансеров и может стать ценным фильтром для определения приоритетности вариантов энхансеров для дальнейшего функционального анализа.
Discussion
Субоптимально аффинные сайты связывания распространены в различных энхансерах, включая Otx-a, svb, ZRS, энхансер IFNβ и многие энхансеры, связанные с заболеваниями, а также энхансеры развития сердца20-22,42. Здесь мы показываем, что однонуклеотидные изменения, незначительно повышающие аффинность связывания, приводят к потере тканевой специфичности и появлению органических фенотипов в конечности мыши и человека. В другом исследовании мы обнаружили, что оптимизирующие сродство SNV в энхансере сердца Ciona приводят к эктопической экспрессии гена FoxF в клетках, не относящихся к сердцу, что вызывает аномальную миграцию клеток и такие серьезные пороки сердца, как два бьющихся сердца42. Наши результаты свидетельствуют о том, что распространенность субоптимальных аффинных сайтов создает уязвимость в геномах, при которой оптимизирующие аффинность SNV могут приводить к эктопической экспрессии GOF и фенотипическим отклонением.
Учитывая наши выводы, для выявления и предсказания случайных вариантов энхансеров необходимо уделять больше внимания низкоаффинным, но высокодегенеративным сайтам. В данном исследовании мы показали, что PBM является высокоэффективным методом измерения аффинности. Хотя связывание in vivo, несомненно, модулируется другими факторами in vivo, такими как меж-белковые взаимодействия, IDR и другие молекулярные взаимодействия, наши результаты показывают, что аффинность связывания транскрипционных факторов является фундаментальной характеристикой, определяющей активность энхансеров и экспрессию генов. Использование такого простого измерения in vitro обеспечивает систематический метод идентификации причинных вариантов энхансеров, который не зависит от специфических измерений в типах клеток. Это позволяет использовать обобщенный подход для выявления причинных вариантов энхансеров GOF, который применим в разных геномах, в типах клеток и даже видах.
Сайт ETS-A представляет собой последовательность длиной 8 п.н., и в этом регионе существует 65 536 возможных комбинаций последовательностей. Из этих комбинаций мы успешно предсказали экспрессию и фенотипы пяти различных последовательностей ETS-A (French 2, Indian 2, Syn 0.25, ETS-A LOF и Syn 0.52) на основе изменений в аффинности связывания. Эти эксперименты демонстрируют возможности механистических правил для предсказания причинных вариантов энхансеров. В будущем мы захотим перейти от простого выявления причинных вариантов энхансеров к предсказанию тяжести и пенетрантности. В сайте ZRS ETS-A большее увеличение сродства приводит к более тяжелым и пенетрантным фенотипам. Вероятно, это справедливо для всех изменений аффинности, происходящих в одном и том же сайте связывания и в одном и том же положении в энхансере, поскольку все эти увеличения аффинности происходят в одном и том же контексте или грамматике. Эффекты SNV, оптимизирующих аффинность, вероятно, модулируются окружающими сайтами связывания, так что варианты в разных позициях энхансера могут иметь разные эффекты, несмотря на одинаковое увеличение аффинности43. Мы видим подтверждение этому в нашем исследовании аффинно-оптимизирующих SNV в сердечном энхансере FoxF у Ciona42. Интеграция понимания аффинно-оптимизирующих SNV и грамматики энхансера улучшит нашу способность предсказывать тяжесть и пенетрантность энхансерных вариантов.
Мы обнаружили, что варианты энхансеров, вызывающие GOF-, но не LOF-энхансерную активность, нарушают развитие. Избыточность энхансеров на нескольких уровнях обеспечивает устойчивость организма. Как правило, несколько энхансеров, известных как избыточные или теневые энхансеры, регулируют один и тот же ген44,45. Другой уровень избыточности энхансеров закодирован внутри энхансеров46,47, примером чего могут служить пять ETS-сайтов (ETS1-ETS5) в ZRS15. Таким образом, потеря одного активаторного сайта, снижение аффинности активаторного сайта или даже потеря всего энхансера может быть компенсирована избыточными сайтами или энхансерами. Напротив, варианты GOF, приводящие к повышению уровня экспрессии или пространственно-временной эктопической экспрессии, сложнее буферизировать и, следовательно, они с большей вероятностью могут повлиять на экспрессию и развитие генов. Примером может служить сайт LOF ETS-A, который не оказывает никакого влияния, в то время как все четыре варианта GOF приводят к эктопической экспрессии генов и нарушают развитие конечностей. Сосредоточение внимания на вариантах, которые приводят к экспрессии GOF, может улучшить нашу способность точно определять причинные варианты энхансеров.
Скопления транскрипционных факторов часто встречаются в непосредственной близости от активных энхансеров. Такие скопления называются хабами и содержат большую концентрацию транскрипционных факторов, которые могут быть разделены по фазе48,49. В такой обстановке изменение одной пары оснований может оказывать столь значительное влияние на экспрессию. На биохимическом уровне мы предполагаем, что едва заметное увеличение сродства позволяет активатору находиться на поверхности более длительное время; это может увеличить вероятность связывания всех необходимых факторов и образования функционального комплекса, способного запустить транскрипцию. Дальнейшие исследования механизмов, с помощью которых один SNV, незначительно повышающий сродство к связыванию, может инициировать активацию транскрипции, могут помочь нам понять движущие силы, лежащие в основе транскрипционного контроля.
Тридцать один SNV в ZRS человека связан с полидактилией. Только семь из них находятся в пределах подтвержденных сайтов связывания; это, вероятно, является результатом вырожденной природы сайтов связывания и нашей слабой аннотации функциональных сайтов связывания в ZRS. Из семи SNV в сайтах связывания три являются SNV, оптимизирующими сродство. Два из этих SNV находятся в сайте ETS-A (French 2 и Indian 2) и один в сайте HOX (Dutch 2). Оптимизирующие сродство к ETS-A SNV вызывают эктопическую экспрессию GOF в переднем зачатке конечности. Стволовые тетраподы были полидактильными с семью или восемью пальцами50. Экспрессия репрессоров в переднем зачатке конечности могла способствовать возникновению пятизначного состояния. Мы предполагаем, что SNV, оптимизирующие сродство к ETS-A, вызывают эктопическую экспрессию в передней части зачатка конечности, поскольку повышение сродства нарушает баланс активаторов и репрессоров, действующих на энхансер в передней части зачатка конечности, а также в силу эволюционной чувствительности.
Миллионы вариантов, которые значительно ассоциированы с фенотипами и заболеваниями, расположены в энхансерах1-3, и функциональное тестирование всех этих вариантов представляет собой серьезную проблему. Эксперименты в стиле MPRA проверяют влияние вариантов энхансеров с помощью репортерных анализов; однако такие анализы имеют ограничения, поскольку они не проверяют варианты в эндогенном локусе, а также в соответствующем клеточном или многоклеточном контексте30,40. Кроме того, MPRA, как правило, выделяют варианты, которые приводят к самым большим и значительным изменениям в экспрессии. Однако они могут не оказывать наибольшего влияния на фенотип. Действительно, варианты French 2 и Indian 2 приводят к эктопической экспрессии всего в нескольких клетках в течение короткого времени, но этого достаточно для формирования дополнительных пальцев. Хотя не все небольшие динамические области эктопической экспрессии будут вызывать фенотип, небольшие изменения во временной и пространственной экспрессии морфогенов, белков сигнальных путей и эффекторов - особенно в типах клеток, в которых они могут изменять идентичность, - вероятно, приведут к дефектам формирования паттерна и фенотипам. Взвешивание степени изменения экспрессии на основе типа гена-мишени и чувствительности клеточного контекста, а также фокусирование на вариантах, которые вызывают GOF, а не LOF экспрессию, может улучшить нашу способность предсказывать варианты энхансеров, которые изменяют фенотипы.
Энхансеры часто классифицируют по нескольким признакам: их способ взаимодействия с промотором; уровень сохранения последовательности; расстояние от целевого промотора; целевой ген; ткань, в которой они активны; и виды, у которых они встречаются. Наши исследования ZRS, сердечного энхансера FoxF, энхансера IFNβ и энхансеров, связанных с заболеваниями, а также анализ eQTL показывают, что использование сайтов с субоптимальной аффинностью для кодирования специфичности энхансера и роль SNV, оптимизирующих аффинность, в экспрессии генов GOF выходят за рамки этих категорий. В более широком смысле, сохранение принципов регуляции в различных энхансерах обеспечивает основу для использования нарушений таких правил для предсказания причинных вариантов, лежащих в основе энхансеропатий.
