Посещений: 
КАРДИОМИОЦИТЫ ИЗ iPSC КАРДИОМИОЦИТЫ ИЗ И ОТ ПАЦИЕНТОВ С КАРДИОМИОПАТИЕЙ
Подходы CRISPR генотерапии
Gene Modulation with CRISPR-based Tools in Human iPSC-Cardiomyocytes • Julie Leann Han & • Emilia Entcheva
 Stem Cell Reviews and Reports (2023)
|
Precise control of gene expression (knock-out, knock-in, knockdown or overexpression) is at the heart of functional genomics - an approach to dissect the contribution of a gene/protein to the system's function. The development of a human in vitro system that can be patient-specific, induced pluripotent stem cells, iPSC, and the ability to obtain various cell types of interest, have empowered human disease modeling and therapeutic development. Scalable tools have been deployed for gene modulation in these cells and derivatives, including pharmacological means, DNA-based RNA interference and standard RNA interference (shRNA/siRNA). The CRISPR/Cas9 gene editing system, borrowed from bacteria and adopted for use in mammalian cells a decade ago, offers cell-specific genetic targeting and versatility. Outside genome editing, more subtle, time-resolved gene modulation is possible by using a catalytically "dead" Cas9 enzyme linked to an effector of gene transcription in combination with a guide RNA. The CRISPRi / CRISPRa (interference/activation) system evolved over the last decade as a scalable technology for performing functional genomics with libraries of gRNAs. Here, we review key developments of these approaches and their deployment in cardiovascular research. We discuss specific use with iPSC-cardiomyocytes and the challenges in further translation of these techniques.

|
Генная модуляция - это изменение гена для механистического изучения биологии или терапевтического вмешательства (рис. 1). Традиционные методы манипулирования экспрессией генов включают в себя малые молекулы, ДНК-связывающие агенты, синтетические олигонуклеотиды или пост-транскрипционные модификации с помощью RNAi. За последнее десятилетие развитие технологий CRISPR расширило набор инструментов для редактирования ДНК или РНК с большей эффективностью и точностью для потенциального применения в сердечно-сосудистой системе [1]. Данный обзор посвящен новым методам модуляции генов с помощью CRISPR, включая те, которые не вызывают необратимых изменений в геноме. Индуцированные плюрипотентные стволовые клетки человека (iPSCs) представляют собой возобновляемый источник труднодоступных типов клеток человека, особенно для изучения кардиологических [2], неврологических [3] и метаболических заболеваний, демонстрируя большой потенциал для разработки/скрининга лекарств, генной терапии и регенеративной медицины. Сочетание этих двух масштабируемых подходов, iPSCs и модуляции генов на основе CRISPR, позволило создать высокопроизводительные методы генетического скрининга для выявления молекулярных основ биологических функций и изучения механизмов заболеваний. Такие инструменты способны ускорить разработку новых целевых терапевтических средств.
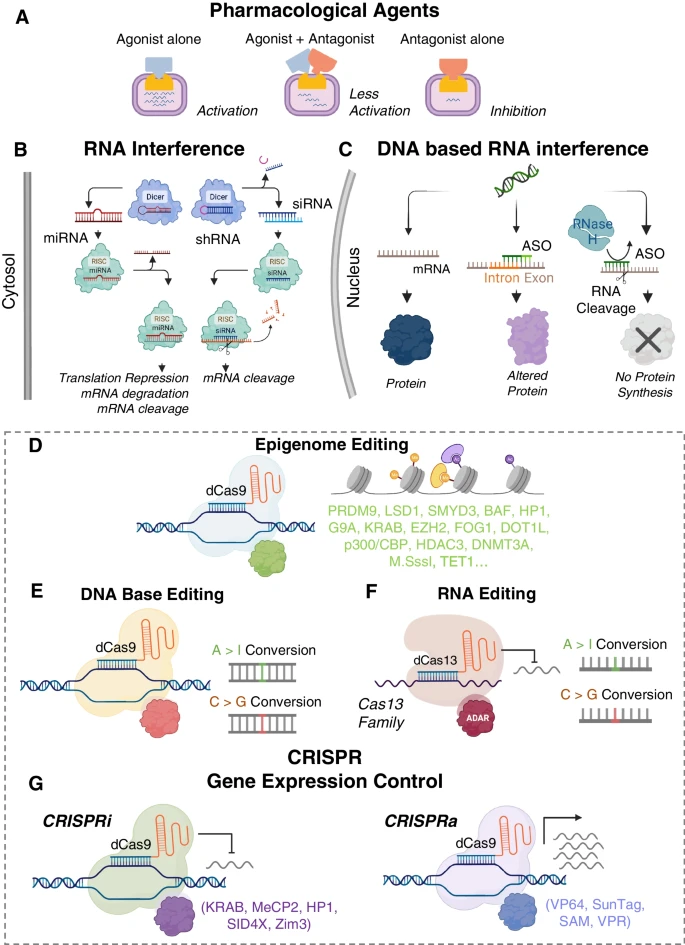 Fig. 1
Approaches for gene modulation. (A) Traditional methods to control gene function involve pharmacological compounds. (B) Targeting miRNAs and shRNA/siRNA in the naturally occurring RNAi pathways has been used to control post-transcriptional gene expression. (C) Basic mechanism of antisense oligonucleotides for targeting protein expression. (D) Epigenome editing with CRISPR. (E) DNA base editing tools introduce single nucleotide edits to the DNA. (F) RNA base editors include cytosine and adenine versions. (G) CRISPR-based approaches for gene activation and interference. Biorender was used for parts of this figure
Classic Methods for Gene Modulation
Transcriptional Modulation by Small Molecules
Традиционные препараты модулируют активность конкретного белка как агонисты (активация), антагонисты (ингибирование) или путем смешанного действия агонист-антагонист, когда они могут обладать как активирующими, так и ингибирующими свойствами, рис. 1А. Малые молекулы могут оказывать краткосрочное или долгосрочное воздействие на экспрессию одного или нескольких генов. Несмотря на последние достижения, специфичность труднодостижима, а вне-целевые взаимодействия являются основным недостатком, поскольку малые молекулы взаимодействуют с непредусмотренными мишенями, что может вызвать фармакологические токсические явления [4].
Традиционные подходы к созданию сердечно-сосудистых препаратов были сосредоточены на доступных мишенях (на поверхности мембраны) и сигнальных каскадах, таких как рецепторы, связанные с G-белками (GPCRs); блокаторы нейрогормонов (катехоламины, ангиотензин, альдостерон); блокаторы ионных каналов; мишени, связанные с патологической нагрузкой (вазодилататоры и диуретики) [5]. iPSC-кардиомиоциты человека экспрессируют ключевые ионные каналы сердца и имеют соответствующий механизм GPCR [6], поэтому они представляют собой более физиологически подходящую альтернативу гетерологичным системам для тестирования новых малых молекул [7]. Полученные от пациентов с определенными мутациями ионных каналов, эти экспериментальные модели предоставляют ценную информацию об эффективности и безопасности вновь разработанных препаратов на доклинической стадии. Например, McKeithan et al. использовали методы высокопроизводительного скрининга в iPSC-CMs человека, полученных от здоровых людей и пациентов с синдромом удлиненного интервала QT типа 3 (LQT3), имеющих мутацию в натриевом канале SCN5a [8]. Путем оптимизации лекарственной химии они получили новые производные mexiletine с лучшей специфичностью и эффективностью для воздействия на поздний натриевый ток, который является виновником LQT3.
Кроме того, протеинкиназы, такие как Ca2+-кальмодулин-зависимая протеинкиназа (CaMK), фосфоинозитид-3-киназа (PI3K), митоген-активируемая протеинкиназа (MAPK), механистическая мишень рапамицина (mTOR), протеинкиназа A (PKA) и протеинкиназа C (PKC) были направлены на фармакологическое лечение сердечных заболеваний, включая сердечную недостаточность, кардиомиопатии, инфаркт миокарда и аритмии [9, 10]. Недавние исследования убедительно демонстрируют полезность iPSC-CMs человека для тестирования ингибиторов MAPK в комбинированной терапии, когда также используются иммунодепрессанты [11] и для крупномасштабного скрининга ингибиторов киназ [12]. Высоко параллельный характер этих анализов и многопараметрические показания позволяют понять механику и могут привести к более надежным предсказаниям действия лекарств.
Другой фармацевтический подход к патологическим кардиомиопатиям заключается в регулировании транскрипции ядерных генов с помощью малых молекул, направленных на эпигенетические модификаторы, например, ацетилтрансферазы гистонов (HATs) и деацетилазы гистонов (HDACs). HATs и HDACs - это главные регуляторы, которые активируют транскрипцию генов путем ацетилирования нуклеосомных гистонов и расслабления структуры хроматина или ингибируют транскрипцию путем деацетилирования нуклеосомных гистонов, соответственно [13]. Например, гидроксамовая кислота трихостатин А (TSA), ингибитор HDAC, является мощным репрессором сердечной гипертрофии и, как было показано, регулирует множество генов в гипертрофическом каскаде [14]. В настоящее время проводится более 500 клинических испытаний ингибиторов HDAC для решения различных задач, в основном связанных с раком [15, 16], а потенциальное кардиотоксическое действие ингибиторов HDAC было проверено в доклинических исследованиях с использованием iPSC-CMs человека [17]. В целом, генная модуляция с помощью малых молекул привлекательна благодаря своему трансляционному потенциалу, однако специфичность действия труднодостижима.
RNA Interference
Ключевым подходом к пост-транскрипционной модуляции генов является РНК-интерференция (RNAi), рис. 1B. РНК-интерференция является зеркальным отражением традиционной лекарственной терапии, поскольку она используется для подавления гена и кодируемого белка определенной мишени [18]. В клетке RNAi происходит естественным образом через микроРНК (миРНК) и другие некодирующие РНК для регуляции экспрессии генов. Ингибирующие РНК могут быть сконструированы так, чтобы имитировать миРНК, малые интерферирующие РНК (siRNAs) или short-hairpin RNA (shRNAs), используя добавки к своим мишеням. siRNAs, химически синтезированные двухцепочечные РНК, осуществляют замалчивание генов путем загрузки на RNA-induced silencing complex (RISC). Это приводит к эндонуклеазному расщеплению пассажирской нити Argonaute-2 и последующей инактивации гена. shRNA представляют собой структуры со стержневой петлей, которые могут быть экспрессированы в векторных системах, таких как плазмиды или вирусные векторы. Они имитируют предшественников миРНК и экспортируются из ядра в цитоплазму с помощью Exportin-5, белка ядерной мембраны, и расщепляются комплексом Dicer в цитоплазме [18]. Образуются дуплексы малых РНК, которые превращаются в зрелые двухцепочечные siRNA и, наконец, приводят к деградации мРНК путем обработки RISC и Argonaute-2.
В отличие от siRNA, которая посттранскрипционно нацелена на конкретный ген, miRNA обладают более широким действием и обычно ингибируют экспрессию генов нескольких мРНК. miRNAs транскрибируются в ядре и дают начало первичной miRNA (pri-miRNA). Pri-miRNA расщепляется с образованием предшественника миРНК и обрабатывается Exportin-5 и комплексом Dicer, как описано выше. После отбрасывания пассажирской нити зрелая одноцепочечная миРНК может целенаправленно воздействовать на мРНК через частичное комплементарное сопряжение оснований, что приводит к замалчиванию целевого гена через репрессию трансляции, деградацию и/или расщепление [19]. Теоретически миРНК имеют более широкое терапевтическое применение, поскольку они могут целенаправленно воздействовать на сложные мультигенные заболевания, например, рак, нейродегенеративные заболевания и сердечно-сосудистые заболевания [20]. В отличие от них, siRNA лучше всего подходят для лечения моногенных заболеваний [21, 22], хотя в некоторых клинических исследованиях сообщалось об использовании siRNA для борьбы с раком и вирусными инфекциями. Человеческие iPSC-CMs являются важной экспериментальной моделью для тестирования siRNAs и miRNAs в терапевтических целях. Например, скрининг в iPSC-CMs привел к открытию миРНК, участвующих в биогенезе предсердного натрийуретического пептида - ключевого биомаркера гипертонии и сердечной недостаточности и потенциальной терапевтической мишени [23].
В 2004 году первый терапевтический препарат на основе siRNA достиг клинических испытаний, в то время как первое клиническое исследование миРНК началось только в 2013 году [24]. Более медленный прогресс в разработке лекарств на основе миРНК может быть связан с неопределенными механизмами действия и специфичностью. В настоящее время существует три препарата siRNA, одобренных FDA, и семь препаратов находятся на поздних стадиях фазы 3 клинических испытаний [25]. В 2018 году patisiran стал первым одобренным FDA терапевтическим препаратом на основе siRNA, который вышел на фармацевтический рынок. Патисиран был разработан для лечения транстиретинового (TTR) амилоидоза, который может привести к тяжелой застойной сердечной недостаточности. Было показано, что он улучшает структуру и функцию сердца у пациентов с кардиомиопатией [25, 26]. Vutrisiran - еще один исследуемый препарат на основе siRNA, нацеленный на TTR, который находится в фазе 3 клинических испытаний для лечения амилоидоза с кардиомиопатией. Это терапия второго поколения с химическими модификациями, введенными для повышения потенции и метаболической стабильности. Inclisiran - это конъюгат siRNA второго поколения, который ингибирует PCSK9 для лечения атеросклеротических сердечно-сосудистых заболеваний путем снижения уровня LDL-C и находится на рассмотрении для утверждения [25]. В целом, RNAi (siRNAs и miRNAs) нашли широкое применение в сердечно-сосудистых исследованиях, в потенциальной терапии сердечно-сосудистыми стволовыми клетками, а также идентификации белок-кодирующих генов и некодирующих РНК в кардиологии [26].
Несмотря на терапевтический потенциал siRNAs и miRNAs, они сталкиваются со значительными проблемами, такими как надлежащая доставка и поглощение целевым участком, низкая биодоступность, быстрое удаление, изменчивость среди типов тканей, активация иммуногенных реакций и деградация нуклеазами, присутствующими в плазме, тканях и цитоплазме [27]. Для улучшения фармакокинетики, фармакодинамики и профиля безопасности были применены химические модификации и оптимизированная доставка [27]. Хотя эти стратегии улучшили стабильность siRNA, в некоторых случаях они также повысили токсичность и снизили подавление генов [28]. siRNA может переносить только ограниченные химические модификации без нарушения активности RISC [29].
DNA-based RNA Interference with Antisense Oligonucleotides
В отличие от siRNA, которые являются двухцепочечными молекулами РНК, антисмысловые олигонуклеотиды (ASOs) представляют собой короткие синтетические одноцепочечные олигомеры ДНК длиной около 10-30 нуклеотидов, которые могут дифференцированно регулировать экспрессию генов, рис. 1С [30]. По сравнению с siRNA, ASO обладают меньшей токсичностью и меньшим внецелевым эффектом при активации иммунной системы хозяина, поскольку они более устойчивы к более широкому спектру возможных химических изменений [31]. ASO могут быть синтезированы либо для понижения регуляции молекулярной мишени, либо для модуляции альтернативного сплайсинга. Чтобы вызвать молчание генов, сконструированная анти-смысловая нить активирует эндонуклеазную активность РНКазы Н, которая расщепляет гибриды РНК-ДНК, значительно снижая трансляцию целевого гена [32]. ASOs также могут быть разработаны для регулирования сплайсинга РНК [30], т.е. удаления интронов из исходного продукта транскрипции ДНК и присоединения белок-кодирующих областей (экзонов) для образования непрерывной молекулы РНК. Альтернативный сплайсинг регулируется для подавления или усиления целевых белков, которые могут производить различные варианты с отличительными функциями. ASOs могут регулировать сплайсинг путем соединения с предшественником мРНК, блокируя связывание факторов сплайсинга, изменяя исходный паттерн сплайсинга и активируя новый сайт сплайсинга, чтобы принудительно включить нужные экзоны [30]. Другие механизмы, управляемые ASO, включают изменение процесса сплайсинга (splice-switching) и стерическое препятствование рибосомальной активности [32].
Существует множество одобренных Управлением по контролю качества пищевых продуктов и лекарственных средств США (FDA) методов лечения с помощью ASOs, и некоторые из них все еще находятся в стадии клинических испытаний. В 1998 году formivirsen стал одним из первых одобренных FDA ASO-препаратов, используемых для лечения цитомегаловируса. В 2013 году mipomersen стал первым ASO, одобренным FDA по сердечно-сосудистым показаниям - лечение семейной гиперхолестеринемии посредством подавления аполипопротеина В, основного фактора сердечно-сосудистого риска [26]. В 2016 году eteplirsen был одобрен для лечения мышечной дистрофии Дюшенна (DMD) [33]. Некоторые недостатки использования ASO в клинике схожи с RNAi в том, что они также подвержены деградации под действием нуклеаз, опасения по поводу равномерной доставки в ткани, несовершенного связывания с целевой мРНК, а также внецелевых эффектов и токсичности [26]. Улучшенные ASOs были разработаны путем введения фосфоротиоатных связей вместо фосфодиэфирных связей между нуклеотидами, образующими основу, для повышения стабильности, увеличения клеточного поглощения и предотвращения деградации [26, 27].
Evolution of the CRISPR Technology
Первоначально идентифицированные как часть естественной системы противовирусной защиты бактерий и архей, CRISPR представляют собой короткие нуклеотидные повторы, которые используются для обнаружения и разрушения ДНК во время инфекции. РНК CRISPR (crRNA) выполняет функцию направления белков Cas на вторгшиеся нуклеиновые кислоты для разрушения нуклеиновой кислоты в ходе врожденного бактериального защитного пути. Ферменты Cas9 в сочетании с последовательностями CRISPR составляют основу технологии CRISPR-Cas9, которая изменила редактирование генома. В 2011 году Charpentier et al. обнаружили, что транс-активирующая CRISPR РНК (tracrRNA) необходима для созревания crRNA в исследованиях на S. pyogenes [34]. Примерно в то же время Siksnys et al. клонировали весь локус CRISPR-Cas9 из S. thermophilus в E. Coli [35]. Они также очистили Cas9 с помощью crRNA и одними из первых охарактеризовали механизм действия Cas9 [36]. Они сообщили, что Cas9 можно направлять на различные сайты-мишени, манипулируя последовательностью crRNA, и определили, что последовательность protospacer adjacent motif (PAM) необходима для первоначального связывания ДНК и расщепления нуклеазой Cas [36]. Похожие результаты были описаны Charpentier и Doudna [37], где они показали, что crRNA и tracRNA могут быть объединены для создания единой синтетической направляющей РНК (gRNA), которая взаимодействует с ДНК-мишенью и с Cas9, что еще больше упрощает технологию. Эти открытия стимулировали усилия по применению технологии для редактирования геномов. Работа группы Zhang's в MIT [38] и группы Church's в Гарварде [39] привела к применению технологии редактирования генов CRISPR в клетках млекопитающих.
Gene Editing with CRISPR-Cas9 Including Prime Editing
Белки Cas, наряду с нуклеазами типа активатора транскрипции (TALEN) и нуклеазами с цинковыми пальцами (ZFN), являются сайт-специфическими нуклеазами, которые позволяют осуществлять генетические модификации путем индукции двухцепочечных разрывов (DSBs) в целевых местах генома. Все они находят широкое применение в экспериментальной биологии и терапевтических целях, но CRISPR/Cas9 превосходит их по точности и специфичности, поскольку для распознавания и расщепления ДНК под руководством РНК требуется всего один белковый домен; в то время как ZFNs и TALENs требуют двух отдельно синтезируемых белковых доменов. В своей классической форме эти инструменты полагаются на активацию двух механизмов репарации ДНК: негомологичного соединения концов (NHEJ) и гомологично направленной репарации (HDR). Путь NHEJ, подверженный большим ошибкам, соединяет фрагментированные концы вместе, что часто приводит к вставкам и делециям (indels), которые приводят к мутациям со сдвигом рамки и последующему нокауту генов. Путь HDR - это точный механизм репарации, который обеспечивает направленную рекомбинацию между шаблоном ДНК-донора и участком разрезанной ДНК для исправления DSB. Следовательно, HDR можно использовать для введения в геном специфических мутаций или трансгенов [40]. Однако многие типы человеческих клеток, включая iPSC-CMs человека, относительно некомпетентны в осуществлении HDR с высокой эффективностью.
Удобно, что CRISPR прайм-редактирование обходит необходимость HDR и было показано эффективным в широком диапазоне типов клеток [41]. В нем используется никаза Cas9 (мутант Cas9, который может вызывать однонитевые "ники"/вырезки), соединенная с модифицированной обратной транскриптазой (чтобы вызвать target-primed reverse transcription (TPRT)) и prime editing gRNA (pegRNA) для внесения новой информации о последовательности в геном в локус-специфический регион без необходимости донорского шаблона [41]. Хотя первоначальная эффективность прайм-редактирования была низкой, модификации улучшили технологию, повысив ее стабильность и улучшив дизайн pegRNA [42]. Прайм-редактирование обычно выполняет более короткие правки длиной около 20 п.н., но может вносить более крупные геномные делеции с обеих сторон целевой ДНК [43]. Прайм-редактирование CRISPR является очень полезной технологией редактирования генома для исследования сложных генетических изменений. Другой подход, основанный на CRISPR, который не вводит DSBs, подобно праймовому редактированию, включает каталитически неактивную dCas9 и рассматривается ниже как часть методов обратимой генной модуляции.
CRISPR/Cas9 уже применялся для исследований in vivo с момента его открытия в 2011 году. S. pyogenes Cas9 (4,1 кбит/с) до сих пор является наиболее широко используемым белком для редактирования генома. Ограничивающим фактором для его применения в клинике была доставка компонентов CRISPR-Cas9 в клетки. Адено-ассоциированные вирусы (AAV) являются наиболее часто используемыми вирусными векторами для клинических и in vivo исследований, поскольку они вызывают слабый иммунный ответ у человека и не интегрируются в геном хозяина. AAVs были одобрены FDA для лечения целого ряда заболеваний. К сожалению, максимальная емкость упаковки AAVs составляет около 4,7 кбит/с, что оставляет мало места для добавления gRNAs или регуляторных факторов для исследований редактирования генов CRISPR in vivo. Одним из методов является прямое введение Cas9 и gRNA белка или мРНК в эмбрионы для развития трансгенных животных, как это, например, сделано для применения в кардиологии в [44-46], Таблица 1. Из-за ограничений, связанных с упаковкой Cas9, gRNAs, упакованных в AAV, они были доставлены в Cas9-экспрессирующие трансгенные мышиные модели [47, 48]. В некоторых исследованиях Cas9 и gRNAs отдельно упаковывались в AAVs для внутривенной доставки [45, 49-53]. Несмотря на ограничение упаковки AAVs, сообщалось об успешной сингулярной доставке Cas9 и gRNA вместе посредством трансдукции AAV для коррекции DMD [54, 55] и коррекции дефектного RyR2 для предотвращения желудочковых аритмий [56]. Альтернативные методы совместной доставки использовали липосомы [57] и аденовирусы [58, 59], несмотря на ограничения клинической трансляции из-за неблагоприятного иммунного ответа и системной токсичности аденовирусов. Для обхода этих ограничений в настоящее время разрабатываются более современные методы использования небольших вариантов Cas9, таких как S. aureus (SaCas9) или S. thermophilus (St1Cas9). Хотя CRISPR/Cas9 является бесценным методом для моделирования заболеваний, его потенциальное использование in vivo требует дальнейшего изучения для преодоления ограничений существующих методов доставки и наблюдаемых внецелевых событий [45, 52, 60]. Например, конструкции AAV-CRISPR вызывали иммуногенный ответ у взрослых мышей, но не у новорожденных животных; глушение Cas9 и gRNAs наблюдалось в течение шести месяцев, а при применении технологии для коррекции мутаций DMD в мышцах были зафиксированы некоторые ограниченные непреднамеренные генетические изменения [52]. Когда система CRISPR-Cas9 была доставлена в iPS-линии, полученные от пациентов, для исправления гетерозиготных мутаций MYBPC3, ответственных за гипертрофическую кардиомиопатию, она вызвала индуцирующие отступы для NHEJ-репарации в более чем половине целевых линий. В отличие от iPS-клеток, когда CRISPR-Cas9 редактирование MYBPC3 проводилось в человеческих эмбрионах, эффективность таргетинга была намного выше, а преобладающим механизмом восстановления был HDR [60].
Table 1 In vivo gene editing with CRISPR/Cas9 - select cardiac applications
Gene Editing with CRISPR/Cas9 in Human iPSC
Редактирование генома в iPSCs может быть использовано для изучения генетических, молекулярных и клеточных механизмов, особенно при кардиологических, нейродегенеративных и метаболических заболеваниях, поскольку иначе трудно получить эти типы клеток и воспроизвести фенотипы заболеваний in vitro. Редактирование на основе CRISPR позволяет создавать изогенные контрольные клетки, в которые вводится или исправляется мутация, связанная с заболеванием, чтобы выявить ее влияние на идентичный генетический фон при моделировании заболевания. В кардиологических приложениях мутации, связанные с болезнью, были исправлены в hiPSC-CMs с помощью CRISPR/Cas9 в качестве потенциального метода лечения кардиомиопатий [2, 64-66]. In vitro Cas9 также применялся для нокаута Nav1.5 для моделирования LQT3 [67] или для нокаута мутанта CACNA1C для моделирования заболевания [68], а также для широкогеномных скринингов на основе iPSC [69-72], Таблица 2. Сочетание этих технологий позволяет количественно оценить роль каждого генетического изменения в контексте заболевания и регенеративной медицины.
Table 2 Cardiac gene editing applications with CRISPR/Cas9 in human iPS cell lines
Использование полученных от пациента hiPSCs позволяет исследовать распространенные и редкие генетические варианты в различных генетических фонах для создания более широких комплексных моделей генетических заболеваний [78, 79]. Это открывает путь к точной медицине и потенциалу для скрининга лекарств, специфичных для пациента, и возможности прогнозировать ответ на клиническое лечение на доклинических стадиях in vitro [7]. Несмотря на потенциал технологии iPSC, существуют некоторые нерешенные проблемы, например, изменчивость линий iPSC, которая может препятствовать точной характеристике генетических вариантов [80]. Получение и дифференцировка iPSC включают процедуры, для которых небольшие вариации на каждом этапе могут значительно повлиять на общий фенотип. Полученные от пациентов iPSC с различным генетическим фоном в значительной степени влияют на функциональность специализированных клеток, полученных из iPSC (например, кардиомиоцитов), также как и негенетические факторы, такие как условия культуры, пассаж и пол [81]. Генетический состав отдельного донора в сочетании с различными протоколами получения iPSC может также влиять на их эпигенетику, тем самым влияя на плюрипотентность и способность к дифференцировке [82]. Кроме того, недавние исследования по достижению высокой экспрессии CRISPR/Cas9 показали, что введение DSBs с помощью Cas9 в iPS-клетки является токсичным с зависимостью от p53 [69]. Это создает большие трудности для успешного и однородного редактирования генома в этих клетках с помощью CRISPR-Cas9 по сравнению с редактированием генома в эмбрионах или других типах клеток [60]. Возможность применения некоторых подходов к генной модуляции в пост-дифференцированных клетках может помочь обойти эти проблемы и по-прежнему использовать изогенные пары для определения того, как конкретный генетический вариант может быть вовлечен в сердечные, нейродегенеративные и метаболические заболевания и развитие.
Gene Modulation with CRISPR-based Methods
CRISPR for Epigenetic Control
Помимо исследований по редактированию генов (knockout/knock-in), технология CRISPR используется для методов генной модуляции без двухцепочечных разрезов в ДНК и может быть развернута в пост-митотических клетках по времени. К ним относятся активация генов (CRISPRa), ингибирование/интерференция генов (CRISPRi), редактирование эпигенома, редактирование оснований ДНК и редактирование оснований РНК, рис. 1D-G. В этих методах используется каталитически мертвая Cas9 (dCas9) с сохраненной способностью нацеливания на сайт. В сочетании с соответствующими эффекторами (транскрипционными факторами) и gRNAs, вместо индукции DSBs, dCas9 может применяться для активации или ингибирования в определенном месте. CRISPRi/a базируются на действие через эпигенетические регуляторы, вовлеченные в метилирование ДНК, ацетилирование гистонов или метилирование гистонов. Поэтому существует значительное совпадение между CRISPRi/a и методами эпигенетической инженерии. Например, Krüppel-associated box (KRAB) для домена CRISPRi индуцирует метилирование гистонов для инактивации генов. И наоборот, эффекторы, используемые в CRISPRa, такие как VP64 [83], VPR [84], Suntag [85] и Synergistic activation mediator (SAM) [86] вызывают эпигенетические изменения, например, ацетилирование гистонов, для активации генов [87-90]. Эти транскрипционные факторы также применялись для исследований эпигенетической инженерии [87, 88, 90]. Эпигенетический контроль - это мощный способ модуляции генов путем внесения химических и топологических изменений в организацию ДНК. Эти факторы вносят вклад в эпигенетическое состояние клетки и модулируют экспрессию генов, судьбу клеток и, в конечном итоге, клеточный фенотип. Многочисленные исследования показали, как эпигенетические модификации могут влиять на развитие и заболевания сердца [1, 16, 91]. Однако до недавнего времени у нас не было инструментов для изучения того, как каждая эпигенетическая особенность может способствовать изменениям в сердечной функции.
Для локус-специфического редактирования хроматиновых меток были использованы ферменты dCas9 для рекрутирования различных эпигенетических эффекторов, рис. 1D. Метилирование по H3K4 приводит к повышению транскрипционной активности, а метилирование H3K9me, H3K27me и H3K79me - к репрессии генов [88]. Ацетилирование H3K27 также было связано с активными промоторами и энхансерами [88]. Недавно Nunez et al. продемонстрировали CRISPRoff - инструмент для программируемой эпигенетической памяти на основе метилирования ДНК, который может вызывать наследственное ингибирование генов [92]. Комбинируя дифференциальные эпигенетические модификаторы и широко нацеленные dCas9s , можно определить, как изменения в механике хроматина в конкретном регионе ДНК влияют на экспрессию генов. Например, dCas9, слитый с гистоновой деметилазой LSD1, может быть использован для специфического определения и целенаправленного воздействия на энхансеры [93]. Эти инструменты помогают определить эпигенетические манипуляции, которые способствуют биологической функции. К сожалению, редактирование хроматина приводит к скромным изменениям экспрессии генов по сравнению с модуляцией генов CRISPRi/a [84]. Таким образом, для достижения долгосрочных, значительных изменений в геноме все еще существует необходимость дальнейшего развития редакторов хроматина.
Gene Modulation by DNA Base Editing
Редактирование оснований - это недавний подход к модуляции генов, и некоторые его варианты являются производными CRISPR/dCas9, рис. 1E. Стратегии редактирования оснований используют специфичность CRISPR, но обходят некоторые ограничения нуклеазы Cas9, а именно: низкую эффективность механизма HDR, необходимость в донорской ДНК, токсичность из-за DSBs [69] и невозможность использования CRISPR/Cas9 для пост-митотических клеток. Были разработаны редакторы оснований, позволяющие осуществлять направленную точечную мутацию одного основания ДНК, не вызывая DSBs и не нуждаясь в донорских шаблонах [94, 95]. Современные методы включают редактор оснований цитозина (катализирует переход C > T на нити с PAM или переход G > A на целевой нити) или редактор оснований аденозина (катализирует переход A > G на нити с PAM или переход T > C на целевой нити), соединенные с dCas9 [1, 95]. Для создания первого редактора оснований Komor et al. использовали фермент цитидиндеаминазы природного происхождения APOBEC1, слитый с dCas9 [94]. Когда gRNA направляет слитый белок APOBEC1-dCas9 (BE1) к целевому сайту, деаминаза преобразует C в урацил (U), который обладает свойствами сопряжения оснований с тимином (T), в пределах примерно пяти нуклеотидов. Далее они модифицировали BE1, чтобы облегчить удаление U из ДНК в клетках и инициировать эксцизионную репарацию оснований (BE2). Затем они устранили несоответствия U:G в пары U:A и T:A (BE3), значительно повысив эффективность редактирования оснований, при этом образовалось менее 1% indels [94].
Поскольку в природе аденозиндезаминазы, действующие на одноцепочечную ДНК, неизвестны, Guadelli et al. трансформировали бактериальный фермент TadA, который естественным образом преобразует A > I/G в РНК, чтобы получить субстрат ДНК для использования в клетках млекопитающих и назвали его ABE7.10 [96]. Более новые версии редакторов оснований, такие как BE4 [97] или ABE8 [98], обладают повышенной эффективностью в клетках млекопитающих, что позволяет проводить скрининг генетических вариантов с редактированием оснований [99] и применять их in vivo [59, 100, 101]. Аденовирусная доставка BE3 была использована для коррекции PCSK9 для лечения атеросклеротических сердечно-сосудистых заболеваний [59, 100, 101], и BE3, как сообщается, обладает более точным редактированием, чем Cas9, без внецелевых событий и хромосомных транслокаций. Компания Verve Therapeutics недавно получила разрешение регулирующих органов на использование этого метода у людей. До этого компания Beam Therapeutics получила разрешение США на начало клинических испытаний терапии редактирования оснований для серповидно-клеточной болезни. Несмотря на эти достижения, поскольку размер spCas9-BE3 (5,1 кб) превышает возможности упаковки AAV, все еще необходимы стратегии по разработке более мелких, функционирующих белков для безопасного внедрения в клинику. Кроме того, деаминаза всегда активна, что усиливает внецелевые эффекты, вызывая побочные правки. Было показано, что в пост-митотических клетках редактирование оснований эффективно лишь на 10%, что все же является улучшением по сравнению с HDR [102].
RNA Base Editing by dCas13
Редактирование на основе РНК использует семейство ферментов Cas13 и его каталитически неактивные аналоги (dCas13) для действия на РНК, а не на ДНК (рис. 1F). Компоненты белка и РНК аналогичны CRISPR/Cas9 и программируются для расщепления РНК или редактирования оснований РНК посредством RNA Editing for Programmable A to I Replacement, version 2 (REPAIRv2) или RNA Editing for Specific C-to-U Exchange (RESCUE) [103, 104]. REPAIRv2 включает неактивный/dead Prevotella sep. P5-125 Cas13b (dPspCas13b) и мутантный домен деаминазы ADAR2, чтобы вызвать редактирование A > I [103]. Путем белковой инженерии и направленной эволюции ADAR2 был слит с dRanCas13b и назван RESCUE, что позволяет осуществлять редактирование C > U [103, 104]. Редактирование РНК имеет ряд преимуществ перед редактированием ДНК, например, обратимость и минимальная цитотоксичность, поскольку действие осуществляется только пост-транскрипционно. РНК-редактирование также может применяться к неделящимся клеткам, например, кардиомиоцитам, поскольку оно не зависит от эндогенных механизмов восстановления, таких как NHEJ и HDR.
Многие заболевания вызваны сплайс-вариантами, которые приводят к мутациям усиления функции, мутациям потери функции или накоплению транскриптов, содержащих повторы, что приводит к образованию аномальных очагов РНК в ядре. Редактирование оснований РНК является удобным подходом для изменения вариантов сплайсинга, нарушения сопряжения оснований РНК-РНК или устранения токсичных РНК в качестве потенциальных стратегий для преодоления болезни. Этот подход может стать полезным инструментом для лечения вирусных инфекций или заболеваний, которые изменяют функцию белка, вызванную трансдукцией сигнала [103]. Ферменты семейства Cas13 имеют небольшие размеры и могут быть упакованы в AAVs, что позволяет использовать их в трансляционной медицине [105].
CRISPRi/CRISPRa
Подобно редактированию эпигенома, большинство технологий CRISPRi и CRISPRa используют подмножество транскрипционных факторов для активации или ингибирования генов, рис. 1G. Эволюция методов CRISPRi/a показана на рис. 2. В клетках бактерий и млекопитающих было замечено, что одна только dCas9 все еще может нацеливаться на сайты транскрипции генов и блокировать транскрипцию без изменения ДНК, что получило название CRISPR-интерференции (CRISPRi) [106]. CRISPRi представляет собой альтернативный или дополнительный подход к нокдауну генов, позволяющий избежать клеточной токсичности и повысить специфичность изменения транскрипции при сохранении генетической структуры [3, 107]. Для повышения эффективности глушения генов с помощью CRISPRi были изучены дополнительные репрессоры транскрипции [106]. Было обнаружено, что dCas9-KRAB оказывает пятикратное подавление по сравнению с двукратным подавлением только dCas9 [108]. Домен KRAB взаимодействует с KAP1, который рекрутирует ингибирующие факторы гетерохроматиновый белок 1 (HP1), гистоновые деацетилазы и SETDB1 для подавления транскрипции [109, 110]. Поскольку подавление, опосредованное dCas9-KRAB, варьирует от гена к гену [108], Carleton et al. исследовали комбинаторные связи между энхансерами и обнаружили, что добавление домена SID к dCas9-KRAB улучшает нокдаун [111]. Домен SID рекрутирует деацетилазы гистонов 1 и 2 (HDAC1/2) и удаляет маркеры ацетилирования гистонов, связанные с активацией [112]. Хотя слияние dCas9-SID было использовано в качестве энхансера [86], предыдущие исследования с помощью TALENS показали, что домен SID может быть использован для репрессии транскрипции [113]. Yeo et al. сконструировали и проверили более эффективный транскрипционный репрессор [114]-MeCP2, который связывается с другим набором транскрипционных регуляторов. Они обнаружили, что KRAB-MeCP2 был наиболее эффективным во всех протестированных мишенях и демонстрировал улучшенную репрессию по сравнению с dCas9-KRAB [114]. Дальнейшие разработки привели к созданию супер CRISPRi, где были слиты два транскрипционных репрессора - белок гетерохроматина 1 (HP1a) и KRAB с белком оболочки MS2, и было сообщено о превосходной репрессии интересующих генов in vivo по сравнению с ранее опубликованным MeCP2 [115]. Белок HP1a содержит хромодомен и CS домен, которые взаимодействуют с метилированным гистоном H3 лизин 9 (H3K9) и H3K9-специфическими гистоновыми метилазами [116]. Совсем недавно Alerasool et al. сообщили о различных доменах KRAB для улучшения подавления и обнаружили, что KRAB-ZIM3 был стабильно более мощным, чем KOX1-KRAB и KOX1-KRAB-MeCP2 [117]. Кроме того, KRAB-ZIM3 меньше, чем KOX1-KRAB-MeCP2, что выгодно для вирусной доставки, и менее чувствителен к выбору gRNA, чем ранее разработанные системы [117]. Коммерчески доступная система CRISPRmod CRISPRi компании Horizon Discovery использует dCas9, слитый с SALL1 и SDS3, для ингибирования транскрипции генов путем рекрутирования белков, участвующих в ремоделировании хроматина и замалчивании генов. Они отметили, что dCas9-SALL1-SDS3 был одинаково специфичен по сравнению с dCas9-KRAB, но был более мощным в репрессии генов-мишеней, основываясь на результатах более раннего исследования [112]. Хотя системы CRISPRi были приняты для одноцелевого и мультиплексного сайленсинга генов [118], эти подходы не всегда последовательны [119] и требуют дальнейшей оптимизации.
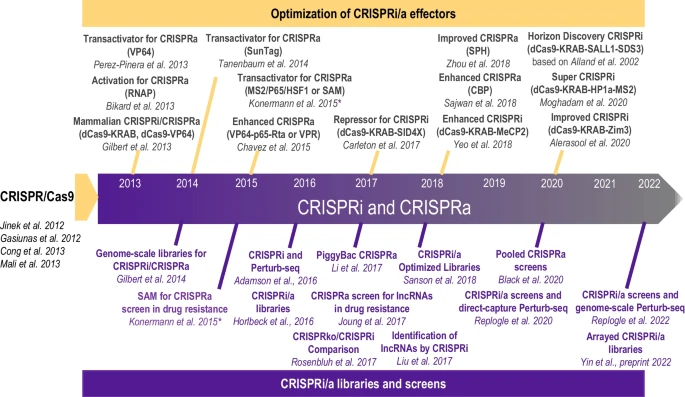 Fig. 2
Timeline of CRISPRa and CRISPRi advancements. Developments of CRISPR-based tools for gene activation (CRISPRa) and inference (CRISPRi), and their expansion into genome-scale gRNA libraries and screens
Слияние dCas с ген-регулирующими белками также используется для активации генов (так называемый CRISPRa), рис. 1G и 2. Методы активации генов были первоначально опубликованы в 2013 году с использованием VP64 [83] и РНК-полимеразы (RNAP) [120]. VP64 является сильным трансактивационным доменом, который рекрутирует HAT p300 и активационные комплексы, вызывая метилирование ДНК, увеличение доступности хроматина и активацию генов [89]. dCas9-VP64 является CRISPRa первого поколения и достигает скромных уровней активации. Эффекторы для CRISPRi, разработанные позже, такие как SunTag [85], SAM [86] и VPR [84], рис. 2, все демонстрируют повышенную активацию генов по сравнению с первоначально разработанным dCas9-VP64 [83] и обеспечивают гибкие альтернативы для дизайна экспериментов. SunTag - это система активации, которая использует каркас из нескольких активаторов VP64 к dCas9 для разделения на параллельные действия транскрипционного механизма, привлекаемого к каждому гену, демонстрируя более сильную активацию с помощью одиночной gRNA по сравнению с dCas9-VP64 [85]. SunTag превосходит активаторы первого поколения, но демонстрирует более низкий уровень активации, чем SAM. SAM использует слитый белок dCas9-VP64 и сконструированные sgRNAs для увеличения транскрипции. Инженерия включает модификацию части gRNAs в аптамеры, нацеленные на MS2 [86], которые затем привлекают дополнительные активационные домены: фактор теплового шока 1 (HSF1) и субъединицу p65 комплекса NF-κB. Было показано, что SAM демонстрирует наиболее эффективные уровни активации для одногенных мишеней. VPR - VP64/p65/Rta [84] был разработан для активации транскрипции с использованием трех мощных эффекторов - VP64, p64 и Rta - слитых с dCas9. Несмотря на более низкую эффективность активации по сравнению с SAM, VPR привлекателен для доставки, поскольку представляет собой однокомпонентную систему. Для мультиплексной регуляции генов SAM, SunTag и VPR показали сходную способность к активации. Появляются новые гибридные методы активации генов, такие как SunTag-p65-HSF1 (SPH) [121], который заменяет домен VP64 в SunTag на активационные домены P65-HSF из SAM. Полученный гибрид обеспечивает двух- или трехкратное повышение эффективности активации по сравнению с SAM, SunTag и VPR. Совсем недавно одна группа разработала эффектор на основе домена HAT (CBP), который также превосходит SAM [122].
Use of CRISPRi/CRISPRa in Cardiac Applications
CRISPRi/a можно использовать для идентификации ключевых генов в развитии и заболевании сердца in vivo и in vitro, Таблица 3. Friedman et al. [123] применили CRISPRi и провели обширный анализ секвенирования РНК в одной клетке во время дифференцировки iPSC-CM, чтобы выявить генные сети для более взрослого фенотипа. Используя индуцибельную систему CRISPRi, Eskildsen et al. [124] определили, что MESP1, критический фактор транскрипции в раннем развитии сердца, также необходим для спецификации сосудистых предшественников. Neiman et al. [125] наблюдали активное участие интегринов (субъединица альфа5) в дифференцировке и сократимости сердечных стволовых клеток, предполагая их роль на ранних стадиях спецификации мезодермы и их подавление при дифференцировке кардиомиоцитов. Schoger et al. [126, 127] создали гомозиготные линии CRISPRi и CRISPRa hiPSC. Важно, что в этих клеточных линиях CRISPRi и CRISPRa не изменили способность клеток дифференцироваться в три зародышевых слоя и производить функциональные iPSC-кардиомиоциты. В рамках исследований, проводимых во время пандемии COVID, Samelson et al. [128] использовали CRISPRi для идентификации бромодомен-содержащего белка 2 (BRD2) как необходимого для транскрипции ангиотензин-превращающего фермента (ACE2) в кардиомиоцитах и, следовательно, потенциально хорошей терапевтической мишени для COVID. Jiang et al. [129] использовали CRISPRa для перепрограммирования фибробластов в клетки-предшественники сердца для имплантации в инфарктные области сердца с целью регенеративной терапии. CRISPRi/a чаще используются в нейроразвивающих приложениях, но они также значительно улучшили использование iPSCs для моделирования сердца in vitro [2].
Table 3 Gene modulation with CRISPRi/CRISPRa in cardiac applications in vitro and in vivo
Методы CRISPRi/a также предоставили новые возможности для изучения патогенеза сердечных заболеваний и разработки более эффективных методов лечения в трудноизучаемой области. Mandegar et al. [130] первыми разработали индуцибельную платформу CRISPRi в iPSCs человека и провели последующее РНК секвенирование, показавшее, что она превосходит CRISPR с активным Cas9, а также обеспечивает обратимую модуляцию генов. Близость к сайту начала транскрипции (TSS) при разработке gRNAs была хорошим предсказателем эффективности. Помимо демонстрации полезности и специфичности CRISPRi нокдауна генов, участвующих в дифференцировке сердечных клеток, и иллюстрации временной генной модуляции экзогенных мишеней (например, датчика кальция GCaMP), они также обнаружили ожидаемые фенотипические последствия (удлинение потенциала действия) CRISPRi снижения калиевого ионного канала HERG в hiPSC-CMs [130]. Limpitikul et al. [131] продемонстрировали новую терапию для персонализированной медицины: CRISPRi в человеческих iPSC-CMs с gRNAs, нацеленными на мутации кальмодулина (CALM), ассоциированные с синдромом удлиненного интервала QT, и скорректировали удлинение потенциала действия из-за избыточного высвобождения кальция. Han et al. [132] разработали масштабируемую платформу CRISPRi для возмущения генов в сочетании с методами определения характеристик на основе оптогенетики (полностью оптическая электрофизиология). Они продемонстрировали корреляционные результаты изменения мРНК с помощью CRISPRi в ключевых ионных каналах сердца и электрофизиологические функциональные эффекты в тех же образцах. В целом, при совместном использовании CRISPRi/a и iPSCs, полученных от пациентов, эти технологии помогают усилиям по персонализированной медицине и лечению, ориентированному на конкретного пациента.
Эти достижения способствовали проведению трансляционных исследований in vivo, Таблица 3. Например, недавно Schoger et al. [133] использовали CRISPRa для активации Mef2d и Klf15, транскрипционных факторов, контролирующих сердечную гипертрофию и гомеостаз, в качестве доказательства внедрения CRISPRi/a для контроля транскрипции эндогенных генов в сердце. Приложения для нейронауки in vivo стали развиваться быстрее. Например, Lau et al. [134] использовали AAVs для системной доставки инструментов CRISPRi/a в мозг мыши для целенаправленного вмешательства и активации эндогенных генов. Colasante et al. [135]. уменьшили судороги путем повышения уровня гена калиевого канала (Kcna1), продемонстрировав подход к лечению эпилепсии на основе CRISPRa. Gemberling et al. [136] разработали модель трансгенной мыши с Cre-индуцибельной CRISPRi/a для контролируемой регуляции целевых генов в печени, Т-клетках, фибробластах и нейронах. Дальнейшие разработки in vivo и потенциальный клинический перевод инструментов CRISPRi/a для использования в кардиологии сталкиваются с теми же проблемами, что и другие методы, связанные с CRISPR, из-за трудностей упаковки и трудностей в достижении эффективной доставки/экспрессии относительно больших конструкций. Оптимизация этих инструментов, включая быстрый подбор gRNA для эффективного ингибирования/активации генов, может ускорить терапевтическое использование in vivo.
CRISPRi/a Screens and Functional Genomics in iPSC-CMs
CRISPR-скрининги являются мощной платформой для геномных и высокопроизводительных генетических исследований с целью изучения генов, путей и механизмов для биологических открытий [137]. По сравнению с традиционными библиотеками RNAi для исследования потери функции, CRISPR и gRNA предоставляют более богатый набор подходов для исследования ингибирования транскрипции, активации, нокаута на большем наборе генов. Исследование до тысячи генов для выявления тех из них, которые важны для биологических путей и потенциальных мишеней для лекарств, возможно в кардиологии, неврологии, метаболических заболеваниях, раке, иммунологии и других областях. На рисунке 2 показано развитие геномных CRISPRi/a-опосредованных скринингов за последнее десятилетие. Gilbert et al. [138] опубликовали первые геномные скрининги для исследований CRISPRi и CRISPRa. Различные усилия были направлены на оптимизацию геномных библиотек gRNA [139-141]. Методы скрининга позволили более эффективно выявлять гены, необходимые для выживания клеток [142], устойчивости к лекарствам [86, 143], упаковки белков [144] и дифференциации iPSC [145-147], рис. 2, табл. 4. Более поздние исследования расширили методы скрининга, включив в них технологии эпигеномного, редактирования оснований и прайм-редактирования [43, 148, 149]. Особенно мощным подходом к изучению вклада отдельных генов в функцию стало сочетание CRISPRi/a с Pertub-seq [144, 150], включая анализ на уровне одной клетки [151] и на уровне генома [152]. Этот метод, характеризующий полный транскриптомный ответ на возмущение одного гена, является перспективным для получения генных регуляторных сетей, GRNs для надежных предсказаний [150].
Table 4 CRISPRi/CRISPRa-based screens using human iPSCs for cardiac applications
CRISPR скрины делятся на два типа: объединенные скрины и скрины с массивами, рис. 3. В сочетании с дифференцированными человеческими клетками, такими как iPSC-CMs, эти скрининги представляют собой специфический для пациента инструмент функциональной геномики, т.е. определения роли каждого гена в биологии изучаемых клеток/тканей. Масштабируемые показания обеспечиваются методами секвенирования следующего поколения (NGS) и другими масштабируемыми технологиями для фенотипической характеристики. Такие интегрированные инструменты ранее не были доступны для исследований человека, и они подают большие надежды в плане информирования и ускорения терапевтических разработок.
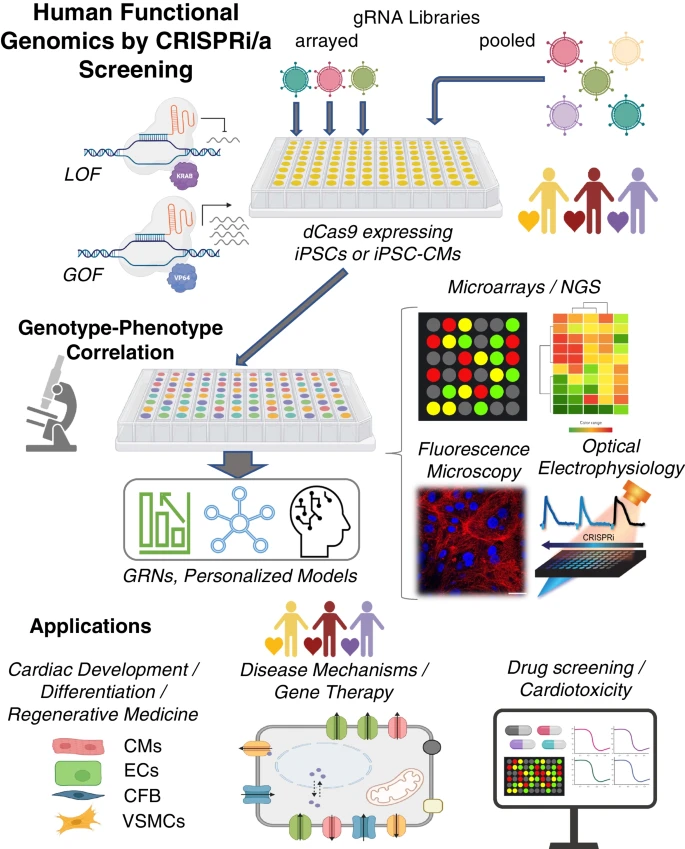 Fig. 3
Human functional genomics by CRISPRi/a. CRISPR-based screening approaches, iPSC technology, and all-optical electrophysiology provide key elements for a high-throughput platform to perturb gene function and analyze respective genomic, protein, and functional changes for biological discovery of cardiac development and disease. Based on combined functional and transcriptomics data, one can build gene regulatory networks, GRNs, using machine learning techniques. The technology and the derived GRN models can be applied to cardiac development, guiding cell differentiation and maturation for regenerative medicine; disease modeling, drug development and cardiotoxicity testing. Biorender was used for parts of this figure
Объединенные скрины CRISPRi/a просты в применении - смешанные gRNAs для всех целевых генов добавляются к образцам сразу. Такие скрининги наиболее подходят для хорошо стратифицированных фенотипов - двух или более типов ответов. Простейшими методами считывания являются анализ жизнеспособности/пролиферации клеток или флуоресцентно-активированная сортировка клеток (FACS). После представления объединенной библиотеки CRISPRi обогащение клеток позволяет проводить механистические исследования выживания и пролиферации клеток, например, объединенный скрининг на основе CRISPRi помог выявить гены, критически важные для роста клеток в iPSCs [145, 156]. Этот метод может быть применен к биологическим результатам, выходящим за рамки выживания, путем сортировки или обогащения клеток по интересующим признакам. Также были созданы клеточные линии, экспрессирующие флуоресцентный белок при активации сигнального пути, которые затем могут быть обработаны для обогащения клеток [157]. Пространственная визуализация в сочетании с маркировкой антителами, малыми молекулами или генетически закодированными репортерами может помочь контролировать клеточную активность отсеянных клеток. Количественное определение мРНК и белка может быть выполнено с помощью флуоресцентной гибридизации in situ (FISH) или проточной цитометрии. Секвенирование РНК одиночных клеток позволяет получить высокоразмерные результаты объединенных скринов CRISPR и уловить биологические фенотипы, которые нелегко измерить с помощью одного маркерного гена.
В отличие от этого, массивы CRISPRi/a скринов обычно представляют одну gRNAs или возмущение для каждой целевой клетки/образца, поэтому активация или ингибирование гена может быть связано со сложными функциональными ответами в разных образцах. Они могут быть соединены с высококонтентными скрининговыми анализами для получения взаимосвязи между возмущением генотипа и более нюансированными клеточными фенотипами. Показания могут быть комплексными - протеомика, метаболомика и функциональная визуализация. Более сложные модели клеток/тканей могут быть объединены с массивами CRISPRi/a скринов, например, 3D органоиды. В целом, объединенные скрининги позволяют совершать открытия, в то время как масс-скрининги лучше подходят для оценки и углубленных механистических исследований. Масс-скрининги gRNAs могут быть объединены с высокопроизводительными полностью оптическими электрофизиологическими исследованиями [158-160] для изучения функциональной геномики развития или заболевания сердца у человека. Fig. 3.
При любом методе генетической модуляции необходимо проверить правильность введения инструментов CRISPR и успешное возмущение генов. Доставка инструментов CRISPR с помощью плазмид, мРНК, белка или лентивирусных векторов может быть сложной в окончательно дифференцированных клетках, таких как кардиомиоциты. Трансдукция с помощью вирусных частиц или методов липофекции и электропорации должна быть оптимизирована для каждого типа клеток и модельного организма. Хотя коммерческие библиотеки gRNAs доступны, панель gRNAs, нацеленных на различные локусы каждого гена, должна быть оценена для определения эффективности возмущения, например, с помощью qPCR, что может быть довольно утомительным, или с помощью более новых методов секвенирования [72]. Кроме того, подтверждение модуляции каждого гена можно провести на белковом уровне с помощью вестерн-блотов или проточной цитометрии. Иногда эти методы не очень хорошо подходят для небольшого объема выборки в высокопроизводительных планшетах, поэтому необходимы новые разработки [161]. В объединенных скринингах клетки могут оцениваться строго на выживаемость и пролиферацию или отбираться под биологическим давлением, таким как лечение лекарственными препаратами или вирусная инфекция, с последующим проведением более глубоких функциональных анализов. Модульные системы, такие как химически- [130, 162, 163] и оптически-индуцируемые [164-166] Cas9/dCas9, позволяют исследовать важные гены с течением времени. Оптический контроль также может обеспечить быструю и точную пространственно-временную модуляцию генов [167]. Создание надежных скринов CRISPR предлагает мощный подход к функциональной биологии.
Conclusions
В последнее десятилетие мы являемся свидетелями слияния нескольких масштабируемых технологий: 1) человеческие клетки iPSC с бесконечной способностью к обновлению; 2) секвенирование нового поколения и транскриптомика отдельных клеток; 3) оптогенетические функциональные анализы; 4) возможности обработки больших данных, мощные и быстрые алгоритмы машинного обучения; и 5) технологии модуляции генов, вдохновленные CRISPR и производные от CRISPR, которые обсуждаются здесь. Полная бесшовная интеграция этих инструментов еще впереди, но они обеспечивают прогресс на пути к функциональной геномике человека, рис. 3. Сочетание этих инструментов позволяет беспрецедентно взглянуть на роль каждого гена в формировании биологических реакций человека в состоянии здоровья и болезни; они могут помочь раскрыть сложные взаимодействия генов на уровне систем, приводящие к определенному фенотипу. Более точные методы генной модуляции, свободные от внецелевых эффектов, обеспечивают критически важный инструмент возмущения для изучения таких взаимосвязей. Достижения в технологии iPSC человека обеспечивают более широкое представительство демографических групп в понимании биологии человека и ее нюансов. Становится возможным тестирование на конкретных пациентах с прямой трансляционной ценностью. Понимание сложных фенотипов, которые сопровождают большинство заболеваний, облегчается благодаря достижениям в транскриптомике и бесконтактным методам функциональной характеристики, выходящим за рамки оценки "живой/мертвый". Огромный объем высоко содержательных данных, полученных в результате слияния этих технологий, обуславливает потребность в более совершенных вычислительных инструментах и алгоритмах обучения. Всесторонняя характеристика реакций может позволить создавать "цифровых двойников" (виртуальные модели) для пациентов в контексте применения лекарств и принятия клинических решений.
Трансляционное воздействие этой конвергенции методов проявляется в информационном обеспечении разработки лекарств, тестирования кардиотоксичности, регенеративной медицины и генотерапии. Кардиология является одной из областей, которые получают наибольшую пользу от этих подходов, благодаря сложной природе функциональных реакций и необходимости создания экспериментальных моделей на человеке. Методы генной модуляции на основе CRISPR уже получили применение in vivo, а некоторые - клиническое воплощение. Ранее не поддававшиеся лечению генетические заболевания могут быть устранены, причем, как можно надеяться, с меньшим количеством побочных эффектов по сравнению с традиционной фармакологией. Общими проблемами для внедрения методов генной модуляции in vitro и in vivo являются эффективные и безопасные методы доставки. Необходимы дальнейшие инвестиции в вирусные и невирусные методы доставки генных модификаций, поскольку они лежат в основе более быстрого внедрения генотерапии в клинику.
|