Calcium signalling and regulation in olfactory neurons
Anna Menini
Current Opinion in Neurobiol. 1999. V.9. P. 419-426
Current Opinion in Neurobiol. 1999. V.9. P. 419-426
Статья сокращена. Перевод В.К. Викуловой
HEK - human embryonic kidney;
IP3 - inositol 1,4,5-triphosphate.
У позвоночных обонятельные сенсорные нейроны локализованы в обонятельном эпителии, выстилающем носовую полость. Это первый этап обоняния. Весь механизм трансдукции - , а именно, превращения воспринятого запаха в электрический сигнал - осуществляется в ресничках этих нейронов. Связывание запаха обонятельными рецепторами в ресничках осуществляется посредством активации G-белком аденилат циклазы, продукции цАМФ, который непосредственно открывает CNG-управляемые канальцы в плазматической мембране. Перенос внутрь потоками Na+ и Ca2+, вместе с Ca2+- активированным током Cl-, деполяризует реснички. (Запахом индуцированный ток может быть измерен с помощью patch-clamping в изолированных обонятельных нейронах.Деполяризация пассивно распространяется по дендриту и телу обонятельного нейрона, запуская потенциал действия, который передается вдоль аксона в обонятельную луковицу (Рис.1)
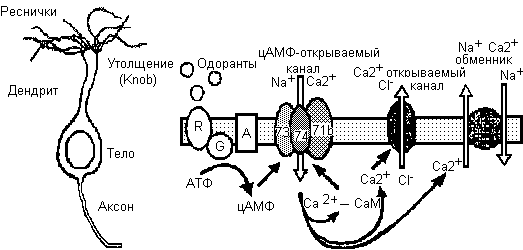
Рис.1 Схема (а) обонятельного сенсорного нейрона и (b) прмнципиальной роли Са2+ в цАМФ- опосредованном пути преобразования в обонятельных ресничках. (а) Обонятельный нейрон состоит из тела, аксона и дендрита с утолщением(knob), из которого выдаются несколько ресничек. Связывание запаха ресничками обусловливает деполяризацию нейрона и создание активного потенциала, который передается вдоль клеточного аксона к обонятельной луковице. Запах связывается с рецепторными белками (R) в мембране ресничек, активизируя тем самым белок G (G). Он в свою очередь стимулирует аденилат циклазу (A), вызывая увеличение концентрации цАМФ. цАМФ непосредственно открывает ионные канальцы в мембране ресничек, вызывая индуцированный запахом направленный внутрь ток, определяемый переносом Na+ и Сa2+. Обонятельные, открываемые с помощью цАМФ, канальцы , как недавно показано, имеют 3 типа субъединиц, называемых здесь 73,74,71b . Увеличение Сa2+ в ресничках вызывает открывание Сa2+-активированных Cl+ канальцев, которые далее деполяризуют клетку. Внутриклеточный Сa2+ связывается с калмодулином (CaM),понижая связывающую чувствительность цАМФ-открываемых канальцев. Сa2+ выталкивается с помощью Na+/Ca2+ обменника.
Второй предполагаемый механизм передачи обоняния связан с образованием инозитол 1,4,5-трифосфата(IP3), но он пока плохо изучен. Более того малое и замедленное возрастания цГМФ (циклического гуанидин-монофосфата), по-видимому, не вовлекается в события первоначального переноса сигнала. Данный вопрос обсуждается в обзоре.
ПОСТУПЛЕНИЕ КАЛЬЦИЯ В ОБОНЯТЕЛЬНЫЕ РЕСНИЧКИ
Измерение внутриклеточного Са2+
Возрастание внутриклетоной концентрации Са2+ в обонятельных нейронах, сопровождающее воздействие запаха измерялось, во-первых, методом imaging, а иногда использовали непрямое маркирование при мониторинге клеток, отвечающих на определенный запах. Исследования успешно решили проблему визуализации сигналов Са2+ в очень тонких обонятельных ресничках (0,1-0,3 m в диаметре). Leinders-Zuffall и др. смогли визуализировать индуцированные запахом преходящие увеличения Са в индивидуальной ресничке и показать путем анализа кинетических и фармакологических свойств увеличение Са в ресничке через CNG каналы.
В ответ на импульс запаха, в ресничке быстро в течение 1 сек. нарастает Са2+ и падает на протяжении 5 сек. Leinders-Zufall и др. также нашли, что увеличение Са в ресничках затем сопровождается увеличением Са в утолщении (knob) дендрита, с запаздыванием в 300 мсек. Затем сигнал из утолщениz дендрита передается в тело нейрона.
Постоянна ли внутриклеточная концентрация Са2+ в ресничках в покое
Из-за очень маленького размера ресничек трудно точно измерить внутриклеточную концентрацию Са2+. Имеющаяся оценка концентрации внутриклеточного свободного Са2+ в покое такова: 40 nM в ресничке и 57nM в утолщениях дендрита.
Субъединицы канала CNG.
Ранее предполагалось, что обонятельные каналы CNG являются гетеротетрамерными комплексами с двумя типами субъединиц; однако, недавно клонирован третий тип субъединицы, являющийся изоформой в фоторецепторах палочках сетчатки (retinal rod?). К сожалению пока не разработана общая номенклатура, я буду следовать номенклатуре of Boenigk et al., и буду называть 73 первую клонированную субъединицу обонятельного канала CNG, 74 вторую клонированную субъединицу, и 71b недавно клонированную субъединицу. В статье Boenigk et al. показал, что нативный обонятельный каналец CNG крысы содержит все три клонированные субъединицы (Рис.1). Все три субъединицы ко-экспрессируются в ресничке обонятельных нейронов крысы. Более того, свойства одного из нативных обонятельных каналов крысы сильно напоминают свойства каналов, состоящих из всех трех субъединиц, экспрессирующихся гетерологически. Стохиометрию обонятельных канальцев CNG, составленных из трех типов субъединиц, остается определить.
Эффект состава субъединиц на ток Ca 2+
В работе, Dzeja et al. исследовано влияние разных субъединиц обонятельных каналов CNG на проникновение Са2+. Показано, что в присутствии миллимолярных концентраций внеклеточного Са2+ относительный поток Са2+ через канальцы, составленные из всех трех субъединиц, на 25% ниже, чем в гомомерных канальцах, составленных только из одной субъединицы 73. Ко-экспрессия субъединиц 73 и 74 незначительно модифицирует относительный поток Са2+ в сравнении с потоком при одной субъединице ?3.
Каковы молекулярные механизмы регуляции поступления Са2+? Хорошо известно, что канальцы CNG разрешают поток Са2+ и в то же время блокируют поток ионов Na +. Показано, что связывающий сайт для Са2+состоит из набора остатков глутамата в гомомерных канальцах CNG фоторецепторных палочек сетчатки и в гомомерных обонятельных канальцах (С Picco, P.Gavazzo, E. Eismann,UB Kaupp, A Menini,неопубл. д.). Замена остатка глютамата глициновым остатком явно уменьшает сродство Са2+ к канальцу, что предопределяет количество Са2+, проникающего в клетку. Следовательно, молекулярной основой изменчивости поступления Cа2+ является состав ее субъединиц, это, вероятно, определяется аминокислотными остатками, отвечающими за связывание Са2+ внутренней стороной поры: критический остаток глутамата в субъединице 73, остаток аспартата в 74 и незаряженный глициновый остаток в 71b.
Какова внеклеточная концентрация Ca2+
Концентрация свободного Ca2+ в слизи эпителии крысы варьирует между 3 и 7 mM (MA Crumling, GH Gold. Soc neurosci Abstr 1998, 29:2099). Эти измерения вместе с измерениями, полученными выше, показали, что в физиологичских условиях Ca2+ составляет, по крайней мере, 80% от общего потока через обонятельные канальцы, открываемые цАМФ.
Белковые токсины: новое средство для изучения канальцев CNG?
Пептидный токсин, который блокирует ток через гомомерные (73) обонятельные CNG канальцы крысы, был выделен из яда змеи Pseudechis australis и был назван псеудехетоксин. Всего 5 nM токсина блокируют 50% CNG-зависимого тока. Остается определить, блокирует ли токсин с высоким сродством и нативные обонятельные канальцы, с тремя типами субъединиц, и блокирует ли другие типы канальцев. Если псеудехетоксин специфичен для CNG канальцев, он может быть очень полезным средством для изучения структуры и физиологии обонятельных канальцев CNG, т.к. уже известные блокаторы не очень специфичны и имеют низкое сродство.
РОЛЬ КАЛЬЦИЯ В ВОЗБУЖДЕНИИ: АКТИВАЦИЯ ПОТОКА ХЛОРИДОВ
Что делает Са2+ на внутренней стороне реснички? Хорошо известно, что Са2+-активированные канальцы Cl- присутствуют в мембране ресничек и что в выделенных обонятельных нейронах запах вызывает рецепторный ток, который имеет два последовательно активируемых компонента. Первый , временно активируемый компонент это преимущественно ток катионов, в основном Са2+, через цАМФ-открываемые канальцы. Когда в ресничке Са2+ накапливается в достаточно высокой концентрации, он открывает Cl- канальцы, продуцируя вторичный поток. В выделенных обонятельных нейронах, 50% кальция участвует в передаче сигналов и регуляции обонятельных нейронов.
До 85% рецепторного тока, индуцированного запахом, может переноситься с помощью Cl-, вытекающего из ресничек, таким образом он вносит свой вклaд в возбуждение клетки. Зависимая от дозы Са2+ активации Cl- канальцев ресничек имеет коэффициент Хилла (Hill) равный 2, это указывает на то, что для открывания одного канальца необходимо кооперативное действие по крайней мере двух ионов Са2+.
Молекулярный состав Са2+-активированных канальцев Cl- еще не определен.
Дальнейшая демонстрация возбуждающей роли потоков Cl-, активированных Са2+, получена в двух работах. Reuter et al.измерил концентрацию Cl- в обонятельной слизистой и в утолщениях дендритов in situ у крыс, используя energy-dispersive X-ray микроанализ. Обнаружено, что концентрация Cl- равна 55 mM в слизи и 69mM - в утолщении дендрита. В другой работе с использованием Cl--чувствительного флуоресцентного окрашивания Nakamura et al. нашел, что средняя концентрация Cl- в обонятельных нейронах тритона примерно 40mM. Следовательно, концентрация Cl- в обонятельных нейронах необычно высока, и, что в физиологических условиях активированные Са2+ канальцы Cl- в обонятельных ресничках выносят Cl- из просвета реснички в слизь.
Следовательно, поступление Са2+ в ресничку предопределяет уровень умножения сигнала через систему обратной положительной связи, которая сцеплена с Ca2+-активированными Cl- канальцами, и клетка деполяризуется без дальнейшего притока в ресничку Са2+.
Kleene исследовал сигнально-шумовые свойства этой усилительной системы и показал, что умножение сигнала посредством Са2+-активированных Cl- канальцев дает очень малое увеличение шума. Единица проведения Са2+-активированных канальцев Cl- оценивается в 0.8 pS и варианса является много меньшей, чем варианса у управляемых цАМФ канальцев. Kleene предполагает, что миллимолярная концентрация Са2+ в слизи оптимизирует вклад двух токов в ресничке (т.е. токов Ca2+ через канальцы, открываемые цАМФ, и Ca2+-активируемых токов Cl-) двумя путями: первый, наружный Ca2+
уменьшает общий первичный поток и его канальный шум, второй, Ca2+ вызывает умножение сигнала посредством низкошумных канальцев Cl-. В результате возникает общий рецепторный ток с двумя, по крайней мере, возможными канальцевыми шумами. Следовательно, поступление Ca2+ продуцирует быстро возрастающее, низкошумовое умножение передачи обонятельного сигнала.
Так как одноканальные потоки через канальцы CNG очень малы, то большая фракция индуцированных запахом токов, переносимых с помощью Ca2+-активированных Cl-канальцев и их биофизические свойства указывают на то, что локальные концентрации Ca2+ внутри реснички высоки. В самом деле, поток Cl- cоставляет половину максимального при цитоплазматической концентрации Ca2+ около 5 microM. Принимая во внимание, что обонятельная ресничка имеет большое отношение поверхности к объему (это примерно цилиндр меньше 0.3 microm в диаметре и 30 microm длиной),то поступление Ca2+ через цАМФ-управляемые канальцы, которые расположены очень плотно на мембране реснички, может легко продуцировать очень высокие уровни Ca2+ внутри ресничек.
РОЛЬ КАЛЬЦИЯ В АДАПТАЦИИ: МОДУЛЯЦИЯ CNG КАНАЛЬЦЕВ
Хорошо известно, что Ca2+ контролирует адаптацию в обонятельных нейронах. Было показано, что принципиальным механизмом регулирования адаптации является Ca2+/калмодулином-опосредованная десенсетизация цАМФ-открываемых канальцев. Авторы впервые исследовали свойства запах- адаптацию в изолированных обонятельных нейронах путем измерения тока в ответ на два коротких импульса-запаха с интервалом в несколько секунд. Ответ на второй импульс был меньше, чем на первый, т.к. приводил к сдвигу в соотношении доза-ответ и к адаптированному к высоким дозам запаха состоянию (Рис 2b).
Сдвиг в соотношении доза-ответ позволяет обонятельному нейрону отвечать на стимулы запаха в очень большом диапазоне концентраций. Подобный сдвиг в доза-ответ соотношении был измерен, когда импульсы запаха давались в присутствии постоянного фона запаха(J Reisert,HR Matthews, abstract in J Physiol 1998, 506P:83: см. замечания в proof).
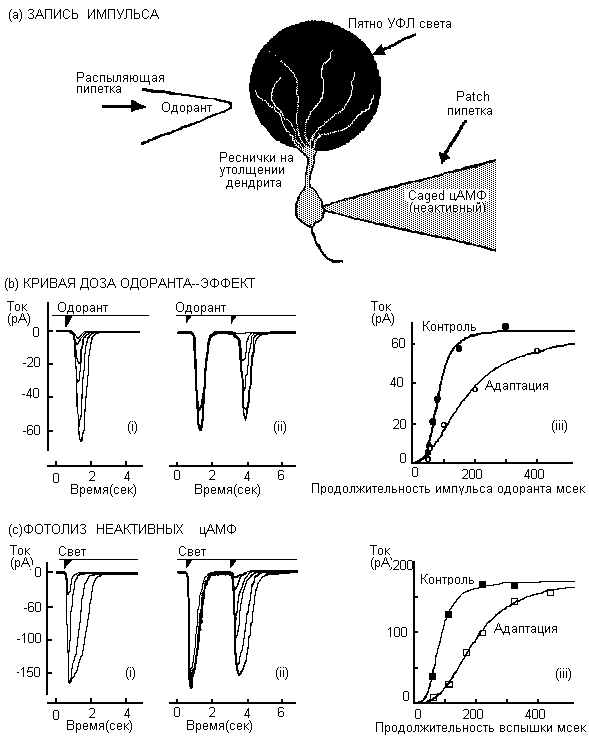
Рис.2 Запись адаптации к запаху в изолированном обонятельном нейроне.
Рис.2 Запись адаптации к запаху в изолированном обонятельном нейроне.(а)Схема, иллюстрирующая экспериментальный метод. Была использована Patch-clamping in voltage-clamp whole-ctll конфигурация для записи тока с изолированного обонятельного нейрона тритона. Напряжение на мембране было -50mV. Обонятельный
стимул доставляли к ресничке путем пипетки-распылителя. Связанное цАМФ, физиологически инертное соединение (но цАМФ можно освободить с помощью УФ света), было включено во внутриклеточный раствор, наполняющий patch пипетку. Через несколько минут после прорыва patch для установления целостности клеточной конфигурации, связанный цАМФ диффундировал внутрь ресничек. Для активизациии цАМФ только внутри области ресничек применяли вспышки УФ.
(b)Дозовая зависимость тока, индуцированного запахом, была измерена путем изменения продолжительности воздействия запаха. (i) Адаптация к обонятельному стимулу определяли на том же обонятельном нейроне при воздействии другого стимула спустя несколько секунд (2,5 сек). (ii) Эксперимент повторяли при сохранении первого обонятельного стимула постоянным, но изменяя продолжительность действия другого стимула. (iii) Взаимосвязь между пиком ответного тока и дозой запаха была схематически изображена и для контрольного и для адаптированного состояний, видно, что при адаптации кривая доза-ответ сдвигается в направлении более высоких доз запаха. (с) Чтобы определить, локализуется ли основной механизм адаптации к запаху до или после продукции цАМФ с помощью аденилат циклазы, в обход индуциуемого запахом ферментного каскада был сделан шунт посредством прямого увеличения концентрации цАМФ внутри реснички путем активизации его при фотолизе. Эксперименты, сходные с описанными в (b) были повторены в отсутствие стимула запахом. (i)Дозовая зависимость фотолизного ответа была сначала измерена в контрольных условиях при изменении продолжительности УФ вспышки, следовательно, при разных количествах освобожденного цАМФ внутри ресничек. (ii)Стимул "адаптирующей" УФ вспышки применяли к той же клетке через несколько секунд (2,5 сек) после первой.
Эксперименты повторяли при сохранении первого УФ стимула постоянным, изменяя продолжительность второй вспышки. (iii)Взаимосвязь между пиком ответного тока и продолжительностью вспышки схематически изображена для контрольного и адаптированного состояний, показывая, что для получения определенного тока в адаптированном состоянии необходима более длительная подолжительномть вспышки, чем в контрольном состоянии. Если адаптация к запаху приписывается снижению продукции цАМФ с помощью индуцированного запахом энзимного каскада, то не должно ожидаться уменьшение тока в ответах адаптированного фотолиза, т.к. вспышки равной продолжительности и интенсивности освобождают равные количества цФМФ. Сравнение между (b) и (с) показывает,что общие свойства адаптации к запаху и фотолизной адаптации очень сходны, показывая, что принципиальные молекулярные механизмы адаптации к запаху действуют после продукции индуцированного запахом цАМФ.Модификация.
Возможные молекулярные механизмы адаптации к запаху включают в себя рецепторы фосфорилирование обонятельных рецепторов, подавление аденилат циклазы, стимуляцию гидролиза цАМФ через активацию фосфодиэстеразы, и модулирование цАМФ-открываемых канальцев. Что бы определить, какой из этих механизмов в интактном выделенном нейроне, Kurahashi and Mеnini исследовали ответное уменьшение при адаптации (Рис. 2a) . Установлено, что ответы, индуцированные цАМФ и запахом имеют сходные свойства адаптации, это указывает на то, что весь процесс адаптации происходит после продукции цАМФ(Fig. 2c). Предполагается, что в процесс гидролиза цАМФ Ca2+-зависимая стимуляция не вовлечена. Следовательно, основной механизм адаптации это Ca2+-зависимая потеря чувтвительности цАМФ-открываемого канальца.
Известно, что Ca2+/калмодулин уменьшает чувствительность обонятельного канальца к цАМФ в 20 раз, предложена модель, основанная на известных влияниях Ca2+/калмодулина на нативный обонятельный цАМФ-открываемые канальцы. Эта модель количественно описывает свойства адаптации к запаху. Модель дает оценку концентрации цитоплазматического Ca2+/калмодулина после первого импульса запаха (около 5 microM).
На молекулярном уровне Ca2+/калмодулин взаимодействует с обонятельным цАМФ-открываемым канальцем путем связывания с коротким сегментом в амино-терминальной области субъединицы 74. Дальнейшие исследования покажут, как взаимодействие Ca2+/калмодулина с другими клонированными субъединицами обонятельного канальца затрагивает проход по нему ионов.
Показано, что Ca2+ модулирует обонятельные цАМФ-открываемые канальцы также и через другой эндогенный неидентифицированный пока фактор, отличный от калмодулина; однако, сравнительная роль калмодулин-зависимого и -независимого механизмов неизучена.
ВЫТАЛКИВАНИЕ КАЛЬЦИЯ ИЗ ОБОНЯТЕЛЬНОЙ РЕСНИЧКИ
Как внутриклеточный Ca2+ возвращается к покойному состоянию после ответа на запах? Можно предположить, что в дендритах, утолщениях дендритов и, возможно, в ресничках есть обменник (exchanger) 3Na+/1Ca2+. Reisert and Matthtws показали, что в обонятельных ресничках лягушки есть Na+-зависимый механизм удаления Ca2+, и, что он отвечает за возвращение внутриклеточного Ca2+ после стимуляции запахом к уровню покоя.
Доказательства Na+-зависимого механизма удаления Ca2+ получены в опытах по замещению ионов. Если Ca2+ выталкивается из реснички с помощью Ca2+ обменника, то замена Na+, омывающего ресничку, другим катионом, таким как холин+ или Li+, известных ингибиторов обменника, должна привести к подавлению истечения Ca2+. В самом деле Reisert and Matthews показали, что ответный ток сильно пролонгирован в отсутствие внешнего Na+, что пролонгированный ток выполняется Ca+-активированными Cl- канальцами, и что для их генерации необходим приток Ca2+ (Рис. 3).Так как Ca2+-активированные Cl- канальцы активируются при концентрации внутриклеточного Ca2+ на уровне микромолей, эти опыты показывают, что Ca2+ не выбрасывается в отсутствии внеклеточного Na+. Следовательно, Na+/ Ca2+ обменник это главный механизм , который возвращает после стимуляции концентрацию Ca2+ в цитоплазме ресничек на основной уровень. Стохиометрия не была измерена в этой работе. Возможно, что и K+ был вовлечен в процесс выталкивания Ca2+ из обонятельных нейронов[60]. Другие механизмы удаления Ca2+ , такие как диффузия внутрь тела клетки или Ca2+-АТФаза[61], вероятно также присутствуют в обонятельном нейроне, но, возможно, количественно они менее значимы. Reisert and Matthews показали, что в отсутствие Na+ восстановления из состояния адаптации после стимуляции не происходит (т.е. когда Na+/Ca2+ не может работать). Следовательно, обменник Na+/Ca2+ отвечает и за восстановление чувствительности после адаптации.
Индуцированные запахом токи генерировали в обонятельном нейроне запахом cineole в растворе Рингера в течение 1 сек. Контролем служила запись при возвращении засасывающей пипетки в раствор Рингера. При экспозиции обонятельного нейрона сразу после стимуляции запахом раствором, содержащим вместо NaCl холин-Cl, был получен Cho+ трек. Известно, что choline+ не может замещать Na+ во многих системах Na+/Ca2+ обменника. В присутствие холина+ наблюдалась реакция пролонгированного тока. Добавление нифлюмовой кислоты, которая блокирует Ca2+-активированный поток Cl-, но не цАМФ-открываемый поток, к раствору холина+ значительно уменьшало амплитуду пролонгированного тока (см. трек Cho++нифлюмовая кислота). Следовательно, большая часть продолжительного тока, наблюдаемого в условиях отсутствия Na+, обусловлена потоком через Ca2+ активированные Cl- канальцы. Эти канальцы активируются концентрациями Ca2+ на уровне микромолей, значит полученные данные показывают, что внутриклеточная концентрация Ca2+ остается высокой во время воздействия раствора с низкой концентрацией Na+, следовательно, подразумевается вовлечение Na+/Ca2+ обменника при восстановлении после ответа на запах. Модификация.
(b)Дозовая зависимость тока, индуцированного запахом, была измерена путем изменения продолжительности воздействия запаха. (i) Адаптация к обонятельному стимулу определяли на том же обонятельном нейроне при воздействии другого стимула спустя несколько секунд (2,5 сек). (ii) Эксперимент повторяли при сохранении первого обонятельного стимула постоянным, но изменяя продолжительность действия другого стимула. (iii) Взаимосвязь между пиком ответного тока и дозой запаха была схематически изображена и для контрольного и для адаптированного состояний, видно, что при адаптации кривая доза-ответ сдвигается в направлении более высоких доз запаха.
(с) Чтобы определить, локализуется ли основной механизм адаптации к запаху до или после продукции цАМФ с помощью аденилат циклазы, в обход индуциуемого запахом ферментного каскада был сделан шунт посредством прямого увеличения концентрации цАМФ внутри реснички путем активизации его при фотолизе. Эксперименты, сходные с описанными в (b) были повторены в отсутствие стимула запахом. (i)Дозовая зависимость фотолизного ответа была сначала измерена в контрольных условиях при изменении продолжительности УФ вспышки, следовательно, при разных количествах освобожденного цАМФ внутри ресничек. (ii)Стимул "адаптирующей" УФ вспышки применяли к той же клетке через несколько секунд (2,5 сек) после первой.
Эксперименты повторяли при сохранении первого УФ стимула постоянным, изменяя продолжительность второй вспышки. (iii)Взаимосвязь между пиком ответного тока и продолжительностью вспышки схематически изображена для контрольного и адаптированного состояний, показывая, что для получения определенного тока в адаптированном состоянии необходима более длительная подолжительномть вспышки, чем в контрольном состоянии. Если адаптация к запаху приписывается снижению продукции цАМФ с помощью индуцированного запахом энзимного каскада, то не должно ожидаться уменьшение тока в ответах адаптированного фотолиза, т.к. вспышки равной продолжительности и интенсивности освобождают равные количества цФМФ. Сравнение между (b) и (с) показывает,что общие свойства адаптации к запаху и фотолизной адаптации очень сходны, показывая, что принципиальные молекулярные механизмы адаптации к запаху действуют после продукции индуцированного запахом цАМФ.Модификация.
Возможные молекулярные механизмы адаптации к запаху включают в себя рецепторы фосфорилирование обонятельных рецепторов, подавление аденилат циклазы, стимуляцию гидролиза цАМФ через активацию фосфодиэстеразы, и модулирование цАМФ-открываемых канальцев. Что бы определить, какой из этих механизмов в интактном выделенном нейроне, Kurahashi and Mеnini исследовали ответное уменьшение при адаптации (Рис. 2a) . Установлено, что ответы, индуцированные цАМФ и запахом имеют сходные свойства адаптации, это указывает на то, что весь процесс адаптации происходит после продукции цАМФ(Fig. 2c). Предполагается, что в процесс гидролиза цАМФ Ca2+-зависимая стимуляция не вовлечена. Следовательно, основной механизм адаптации это Ca2+-зависимая потеря чувтвительности цАМФ-открываемого канальца.
Известно, что Ca2+/калмодулин уменьшает чувствительность обонятельного канальца к цАМФ в 20 раз, предложена модель, основанная на известных влияниях Ca2+/калмодулина на нативный обонятельный цАМФ-открываемые канальцы. Эта модель количественно описывает свойства адаптации к запаху. Модель дает оценку концентрации цитоплазматического Ca2+/калмодулина после первого импульса запаха (около 5 microM).
На молекулярном уровне Ca2+/калмодулин взаимодействует с обонятельным цАМФ-открываемым канальцем путем связывания с коротким сегментом в амино-терминальной области субъединицы 74. Дальнейшие исследования покажут, как взаимодействие Ca2+/калмодулина с другими клонированными субъединицами обонятельного канальца затрагивает проход по нему ионов.
Показано, что Ca2+ модулирует обонятельные цАМФ-открываемые канальцы также и через другой эндогенный неидентифицированный пока фактор, отличный от калмодулина; однако, сравнительная роль калмодулин-зависимого и -независимого механизмов неизучена.
ВЫТАЛКИВАНИЕ КАЛЬЦИЯ ИЗ ОБОНЯТЕЛЬНОЙ РЕСНИЧКИ
Как внутриклеточный Ca2+ возвращается к покойному состоянию после ответа на запах? Можно предположить, что в дендритах, утолщениях дендритов и, возможно, в ресничках есть обменник (exchanger) 3Na+/1Ca2+. Reisert and Matthtws показали, что в обонятельных ресничках лягушки есть Na+-зависимый механизм удаления Ca2+, и, что он отвечает за возвращение внутриклеточного Ca2+ после стимуляции запахом к уровню покоя.
Доказательства Na+-зависимого механизма удаления Ca2+ получены в опытах по замещению ионов. Если Ca2+ выталкивается из реснички с помощью Ca2+ обменника, то замена Na+, омывающего ресничку, другим катионом, таким как холин+ или Li+, известных ингибиторов обменника, должна привести к подавлению истечения Ca2+. В самом деле Reisert and Matthews показали, что ответный ток сильно пролонгирован в отсутствие внешнего Na+, что пролонгированный ток выполняется Ca+-активированными Cl- канальцами, и что для их генерации необходим приток Ca2+ (Рис. 3).Так как Ca2+-активированные Cl- канальцы активируются при концентрации внутриклеточного Ca2+ на уровне микромолей, эти опыты показывают, что Ca2+ не выбрасывается в отсутствии внеклеточного Na+. Следовательно, Na+/ Ca2+ обменник это главный механизм , который возвращает после стимуляции концентрацию Ca2+ в цитоплазме ресничек на основной уровень. Стохиометрия не была измерена в этой работе. Возможно, что и K+ был вовлечен в процесс выталкивания Ca2+ из обонятельных нейронов[60]. Другие механизмы удаления Ca2+ , такие как диффузия внутрь тела клетки или Ca2+-АТФаза[61], вероятно также присутствуют в обонятельном нейроне, но, возможно, количественно они менее значимы. Reisert and Matthews показали, что в отсутствие Na+ восстановления из состояния адаптации после стимуляции не происходит (т.е. когда Na+/Ca2+ не может работать). Следовательно, обменник Na+/Ca2+ отвечает и за восстановление чувствительности после адаптации.
Индуцированные запахом токи генерировали в обонятельном нейроне запахом cineole в растворе Рингера в течение 1 сек. Контролем служила запись при возвращении засасывающей пипетки в раствор Рингера. При экспозиции обонятельного нейрона сразу после стимуляции запахом раствором, содержащим вместо NaCl холин-Cl, был получен Cho+ трек. Известно, что choline+ не может замещать Na+ во многих системах Na+/Ca2+ обменника. В присутствие холина+ наблюдалась реакция пролонгированного тока. Добавление нифлюмовой кислоты, которая блокирует Ca2+-активированный поток Cl-, но не цАМФ-открываемый поток, к раствору холина+ значительно уменьшало амплитуду пролонгированного тока (см. трек Cho++нифлюмовая кислота). Следовательно, большая часть продолжительного тока, наблюдаемого в условиях отсутствия Na+, обусловлена потоком через Ca2+ активированные Cl- канальцы. Эти канальцы активируются концентрациями Ca2+ на уровне микромолей, значит полученные данные показывают, что внутриклеточная концентрация Ca2+ остается высокой во время воздействия раствора с низкой концентрацией Na+, следовательно, подразумевается вовлечение Na+/Ca2+ обменника при восстановлении после ответа на запах. Модификация.