ПОДУШКИ ЭНДОКАРДИАЛЬНЫЕ
Когда образуется сердечная трубка, то кардиальный гель накапливается в атриовентикулярном канале и тракте оттока. Эти области скопления матрикса (примордии подушек) регионально ограничены не только вдоль продольной оси, но и по отношению к поперечным осям сердечной трубки (Eisenberg , Markwald, 1995). В атриовентрикулярном канале подушек вздутия распространяются от дорсального и вентрального квадрантов и от правой и левой частей в конусе вентрикулярного тракта оттока.
Предопределение местоположения эндокардиальных подушек
Так как сердечный гель является в принципе утолщенной миокардиальной базальной мембраной, то локализация частиц адхероновв этом матриксе указывет на то, что они продуцированы миокардом. Эти частицы состоят из 30 и 60 нм субчастиц. Показано (Eisenberg , Markwald, 1995), что частицы содержат боле 10 белков, включаяфибронектин, трансферин и новый белок ES/130 . Эти частицы (адхероны) переносятся в эндокард через кардиальный гель. При этом в тракте оттока и АВ области происходит: гипертрофия эндотелиальных клеток в основном за счет увеличения системы вакуолей ( Гольджи и грубого эндоплазматического ретикулема); потеря межклеточных контактов; способность клеток перемещаться в плоскости эпителия; поляризация Гольджи в будущем конце перемещения трансформированных клеток; образование миграторного аппарата (филиподии и псевдоподии); инвазия в соседний кардиальный гель. Активированнвые эндокардиальные и инвазирующие мезенхимные клетки экспрессируют проколлаген типа I, тенасцин-С, GalTase, урокиназу, коллагеназу, Мох-1, Msx-1, фибриллин-2 (JB3) и гладкомышечный альфа-актин.
Молекулы NCAM экспрессируются во всем эндокраде до образования подушек. Когда эндотелиальные клетки активированы к трансформации и начинают в подушки-формирующих областях отсоединяться от эндотелиального слоя, уровеньNCAM на их поврехности резко снижается как прелюдия к их миграции в кардиальный гель. Когда экспрессия NCAM подавляется, эндотелиальные клетки начинают экспрессировать молекулы адгезии с субстратом(substrate adhesion molecule (SAM)) цитотактин, известный также как тенасцин,. Эти молекулы, которые временно экспрессируются клетками подушек, по-видимому, используются во время инициальной фазы мезенхимной трансфрмации. Тенасцин в других клеточных системах разрушает адгезию между клетками и субстратом, способствуя тем миграции клеток в матриксе. Другой класс молекул, необходимый для миграции мезенхимных клеток - серин протеазы, которые секретируются клетками подушек, когда они трансформируются в мезенхимные.
Позиционирование (предопределение положения) подушек м.б. причинно связано с образованием петли первичной сердечной трубкой. Свет на молекулярный механизм позиционирования подушек может быть пролит с помощью модели развития сердца, которая рассматривает сердечную трубку, как сегментированную вдоль продольной оси.
В сердечной трубке детерминируются две популяции эндокардиальных клеток в сегменте тракта оттока и атриовентрикулярного канала, которые в ответ на сигналы, продуцируемые миокардиальными клетками, трансформируются в мезенхимные клетки (эпителиально-мезенхимная трансформация), которые мигрируют в подлежащий матричный кардиальный гель, образуя ткань подушек.
Целлюляризация (заполнение клетками) подушек
Эндокардиальныe подушки (утолщенные участки кардиального геля)в атриовентрикулярном канале и тракте оттока, по-видимому , несколько отличаются по клеточному составу. В подушках тракта оттока клетки происходят не только из эндотелиальных клеток , но и из клеток нервного гребня из области 3-6 глоточных дуг. Это показано для эмбрионов мыши на стадии 6 сомитов (McMahon et al.,1992). Этот вклад клеток нервного гребня зависит от активности генов Wnt-1,Wnt-3a.
Клетки атриовентрикулярных подушек возникают только из эндокардиальных эндотелиальных клеток, которые трансформируются в мезенхимные. Так как кардиальный гель состоит из молекул матрикса, продуцируемых первоначально миокардом, то наиболее вероятно, что миокард играет важную роль в регуляции трансформации эндотелиальных клеток (Рис.1)
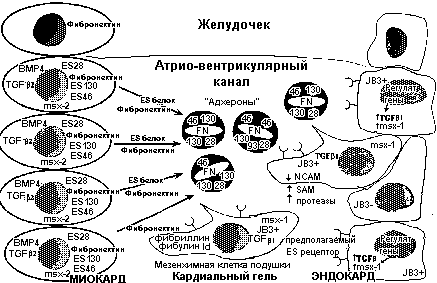
Рис.1. Молекулярные события, участвующие в трансформации эндотелиальных клеток в мезенхиму подушек.
Fig.1. Molecular events, participating in transformation of the endothelial cells in mesenchime of the cushions
Fig.1. Molecular events, participating in transformation of the endothelial cells in mesenchime of the cushions
Миокард примитивной сердечной трубки обнаруживает сегментный паттерн экспрессии молекул. Молекулы, экспрессирующиеся уникально в атриовентрикулярном канале и вентрикулярном тракте оттока, включают TGF-β2, BMP-4, msx-2 и ES белки. Эти последние варьируют в размерах от 28 до 130 кД, они формируют комплексы с фибронектином и ,по-видимому, регулируют трансформацию эндотелиальных клеток. Трансформирующиеся клетки увеличивают экспрессию TGF-β и гомеобоксного гена msx-1. Клетки трансформированные в мезенхимные экспрессируют HLH белок Id и начинают продуцировать белки внеклеточного матрикса фибулин и фибриллин.
Предполагается, что экспрессияTGFβ3 является эндокардиальным ответом на миокардиальный сигнал к трансформации.
TGFβ3 экспрессируется в примитивной сердечной трубке со ст. 8-9. В начале такой трансформации во время кардиогенеза у эмбрионов кур гомодимерный пептид TGFβ3 экспрессируется в эндотелиальных клетках трансформирующихся областей (тракта оттока и АВ области) в трансформирующихся эпителиальных и инвазирующих мезенхимных клетках, тогда как ВМР-2 экспрессируется в соседнем миокарде. Гетеропептидв ВМР2/7 и ВМР4/7 обладают большей биологической активностью, чем гомодимеры в формировании подушек. Низкий уровень экспрессии в миокарде обнаружен и для TGFβ3. TGFβ2 обнаруживается как в эндотелии, так и миокраде перед и после ЭТМ.
время мышинного кардиогенеза мРНК TGFβ1 первоначально экспрессируется во всем эндокарде, а затем ограничивается лишь трактом оттока и АВ областью. TGFβ2 мРНК миокарде тракта оттока и АВ области регионально ограниченным способом, а TGFβ3 мРНК обнаруживатся только после образования подушек.
Миокардиальный ВМР-2 действует синергично с TGFβ3 в инициации эндотелиально-мезенхимной трансформации. Латентные TGFβ комплексы активируются в результате протеолиза и высвобождаются зрелые TGFβ, которые затем соединяются со своими рецепторыми комплексами. ВМР2 усиливает индуцируемые TGFβ инициальные фенотипические изменения, савязанные с ЭТМ. ВМР может менять связываение TGFβ с его рецепторами. Это один из миокардиальных сигналов индуцирующих ЭТМ.
Экспрессия TGF-β эндокардом находится также под регуляцией ES белков. Описан новый белок, GbHLH1.45, член семейства HLH транскрипционных факторов. Он экспрессируется в подушках с начала их целлюляризации и поддерживается на высоком уровне в течение всего развития клапанов. Выявлется перекрывающийся паттерн его экспрессии с другим HLH белом, Id. У него отсутствует ДНК-связывающая область и он оказывает свое влияние на дифференцировку, образуя гетеродимеры с другими HLH белками. Оба GbHLH1.4 и Id экспрессируются в ранних подушках.
Важную роль в этом процессе играет эндоглин.
Важную роль в этом процессе играет эндоглин.
У эмбрионов перепела на ранних стадиях развития (12), т.е. еще до инициации эпителиально-мезенхимной трансформации экспрессируются высокие уровни мРНК для ММР-2. ММР-2 постоянно экспрессируется в популяции этих клеток, т.е. экспрессия не зависит от миокардиальных сигналов, инициирующих процесс трансформации. Однако миокардиальные сигналы обусловливают переход этого фермента в активную форму (66 кДа)на стадиях 18-24, что соответствует времени эпителиально-мезенхимной трансформации и выселению клеток в кардиальный гель.Клетки гипертрофируются их слипчивость друг с другом уменьшается и в них активируются специфические гены, способствующие миграции мезенхимных клеток в матрикс сердечного геля.
В это же время происходит и активация внеклеточной серин-протеазы, урокиназы. Ингибирование связывания с клеточной поверхностью или синтеза урокиназы нарушает двигательное поведение мезенхимных клеток in vitro. Таким образом, урокиназа и ММР-2 обычно экспрессируются на поверхности сердечных мезенхимных клеток, способствуя их миграции и инвазии (Alexander at al., 1997).
На самых ранних из изученных стадий развития мыши(Е10-Е12) обнаруживались различия в мечении поуглерод ангидразе II (Carbonic anhydrase II). Иммуноокрашенные клетки были обильны в атриовентрикулярных подушках, менее многочислены в проксимальных частях конотрункального гребня и редки в дистальной его части. С Е13 окрашивание более регулярно вдоль конотрункального гребня. Между Е13-Е116 сигнал обнаруживался в определенных областях миокарда: передней и левой стенке левого желудочка. На Е18 и Е20 мечение обнаруживалось только в некоторых эндотелиальных клетках коронарных сосудов в частности в межжелудочковой перегородке. Паттерн экспрессии carbonic anhydrase II в активированных эндотелиальных клетках и мезенхимных клетках кардиальных подушек, происходящих из эндотелия, указывает на то, что этот фермент может служить маркером для этой субпопуляции эндотелиальных клеток и их производных с активным поведением (инвазия, миграция, пролиферация)(Vuillemin & Pexieder, 1997).